|
Founded 1871
A Journal of Scottish
Natural History
Editorial
Committee:
J.A. Gibson
John Hamilton
John C. Smyth
A. Rodger Waterston
Reproduced with the permission
of
THE SCOTTISH NATURAL HISTORY LIBRARY
Foremount House Kilbarchan,
Renfrewshire PA10 2EZ
Copyright:
May be used for private research only
Due
to the size of this text file, the many figures
and graphics are individually available
Simply click the underlined links.
The Scottish Naturalist, now published by
the Scottish Natural History Library, is an
independent journal primarily devoted to the
study of Scottish natural history. It was
founded in 1871 by Dr. F. Buchanan White,
of Perthshire, and in 1988 completed one hundred
years of publication. For a summary of the
record of publication, see the inside back
cover.
Although the journal's main interests have always
centred on the history and distribution of
Scottish fauna and flora, it is prepared to
publish contributions on the many aspects
of Scottish natural science embraced by its
title, including Zoology, Botany, Geology,
History, Geography, Archaeology, and the Environment.
All
papers and notes for publication, or books
for review, should be sent to the Editors
at the Scottish Natural History Library, Foremount
House, Kilbarchan, Renfrewshire PA10 2EZ.
Contributions should be clearly written; whenever
possible they should be typed, double‑spaced,
on one side of the paper, with adequate margins,
and should try to conform to the general style
and arrangement of papers and notes in the
current number of the journal. Maps, diagrams
and graphs should be drawn in black ink on
white unlined paper. Photographs should be
on glossy paper. Proofs of all contributions
will be sent to authors and should be returned
without delay.
Authors of papers, but not of short notes, will
receive thirty reprints in covers free of
charge. Additional reprints may be ordered,
at cost, when the proofs are returned.
The Scottish Naturalist is usually published
three times a year. The standard annual subscription
is BP25.00, which should be sent to the Editors
at the Library address. Members of recognised
natural history organisations, however, can
receive the Scottish Naturalist at
a greatly reduced subscription; for details
apply to the Editors.
THE SCOTTISH
NATURALIST
Founded 1871
A Journal of Scottish Natural
History
With which is incorporated The
Annals of Scottish Natural History
and The Western Naturalist
Record
of Publication
The Scottish Naturalist
and Journal of the Perthshire
Society of Natural Science
1871
The Scottish Naturalist
1872-1891
The
Annals of Scottish Natural History 1892-1911
The Scottish Naturalist
191-1939, 1948-1957, 1961-1964
The Western Naturalist
1972-1982
The Scottish Naturalist
1983-date
Published by The Scottish
Natural History Library
LOCH NESS HABITATS OBSERVED BY SONAR
AND UNDERWATER TELEVISION
By
ADRIAN J. SHINE and DAVID
S. MARTIN
Loch
Ness and Morar Project
International Society of
Cryptozoology
Society for the History
of Natural History
Symposium on
The Loch Ness Monster
Royal Museum of Scotland,
Chambers Street, Edinburgh
25th July 1987
Reprinted from The Scottish Naturalist
1988, pages
111-199
LOCH NESS HABITATS OBSERVED
BY SONAR AND UNDERWATER
TELEVISION
By ADRIAN J. SHINE and
DAVID S. MARTIN
Loch Ness and Morar Project
Contents
| Part A: LOCH NESS |
|
| 1. Morphometry. |
|
| Basin Form |
112 |
| History and Sediments |
115 |
2. Thermal Structure. |
|
| Stratification |
118 |
| Physical Studies |
118 |
| Factors affecting Sonar |
119 |
| The Record |
121 |
| Possible Errors |
123 |
| Shear |
126 |
| Conclusion |
128 |
3.Biology. |
137 |
| Phytoplankton |
137 |
| Zooplankton |
137 |
| Littoral and Sub-littoral
Benthos |
140 |
| Profundal Fauna |
144 |
Fish: |
|
|
Littoral Zone
|
149 |
|
Profundal Zone
|
151 |
|
Pelagic Zone
|
154 |
Part B: THE MYSTERY |
|
| 1. The Controversy |
|
| Introduction |
160 |
Biological Considerations |
|
| Habitats |
161 |
| The Candidates |
162 |
Physical Factors and
Inanimate Explanations |
|
| Optical Effects |
163 |
| Submerged Logs |
167 |
| Gothic Revision |
167 |
| Floating Logs |
170 |
2. The Sonar Contribution |
|
| Previous Records |
171 |
Deep Water Contacts |
174 |
| Strength |
177 |
| Depth |
178 |
| Movement |
181 |
| Fish Shoals |
182 |
| Boat Wakes |
182 |
| Thermal Effects |
182 |
| Side Echoes |
182 |
| Tethered Debris |
185 |
Operation Deepscan Supplement |
185 |
Conclusions |
191 |
PART C: EQUIPMENT |
|
| Underwater Photography |
192 |
| Echo-sounders |
193 |
| Scanning Sonars |
193 |
| Temperature probes |
194 |
| Vessels |
194 |
| Additional Equipment |
194 |
PART D : REFERENCE |
|
| Summary |
194 |
| Acknowledgements |
195 |
| References |
196 |
| |
|
Part A: LOCH NESS
1. MORPHOMETRY
Basin Form
Loch, Ness is
the greatest body of fresh water in the British
Isles, having a volume (74.52m3
x 108) in excess of all the lakes
and reservoirs of England and Wales combined
(Smith and Lyle, 1979). It is a glaciated
tectonic lake extending for 35km within the
north eastern section of the Great Glen. The
remarkably regular basin has a mean width
of about 1.5km, with steep sides sloping to
a flat bed, interrupted only by a rise opposite
the Foyers River, which divides the two deep
basins of 220m depth. The maximum depth of
230m is considerably deeper than the seas
around our shores, and places Loch Ness second
only to Loch Morar (310m) among British lakes.
Fault line origins, however, give Loch Ness
the greatest mean depth, of 132m. The catchment
area is 1,775km2, mostly of hard
rock yielding few nutrients.The loch is
drained by the River Ness to the north. Figure
1a
(8K) maps the main river inputs.
The above depths
were determined by Sir John Murray's Bathymetrical
Survey of the Scottish Fresh‑Water Lochs
(Murray and Pullar, 1903-08, 1908a and
1910) using a wire sounding machine, but they
were disputed by a sonar depth of 297m
recorded by Vickers in 1969 (Eastaugh, 1970).
During the 1980s, careful searches of the
reported area, a quarter mile south of Urquhart
Castle, were made by the Loch Ness and Morar
Project using a Kelvin Hughes M.S.48 hydrographic
echo‑sounder, calibrated on site in
accordance with survey practice, but no depths
much in excess of 220m were found.
Figure
1b
(18K) shows a characteristic echo profile
of the loch and contrasts its trench-like
profile with that of Loch Morar. Time advances
left to right on the record and it is important
to note the compression of vertical scale
induced by boat speed. The profile illustrated
in Figure
2a (19K) results from a faster speed
and also serves to illustrate 'side echoes',
which in the past have been interpreted as
extensions of the side walls continuing down
beneath loose sediment for hundreds of metres.
As the conical sonar beam profiles 'down'
a steep slope it registers not vertical depth
but the range of the nearest portion of slope,
which is to the side and therefore shallower
than true depth. As the loch bed begins to
register and the boat draws clear of the wall,
the beam's outer edges and side lobes still
return echoes from the side, which now appear
at a greater depth, thus giving rise to the
above impression. A sonar frequency of about
10kHz or less is required for true sediment
penetration. Underwater television pictures
in Figure
2b (15K) of the steep rocky walls,
contrast with the flat silt of the abyssal
plain.
There is no foundation for speculations concerning
ancient constructions, since there is no evidence
that the loch level was ever much lower than
it is today; nor of submerged caverns, subterranean
connections and the like, since there is no
limestone in the area.
Great advances have been made in hydrographic sonars.
In September 1987 Simrad U.K. Ltd extended
a collaboration while their demonstration
vessel R/Y Simson Echo was in
the loch. The vessel mounted a multibeam or
'swath' sounder (E.M. 100). Data is recorded
continuously from 32 beams, which form a fan
athwartships. This greatly reduces the number
of survey lines necessary and hence the time
required.
On 7th September 1987 a survey was made of Urquhart
Bay. A graphic, constructed in Figure
3 (13K), serves to show the bay contours sloping away from the river mouths and
then dropping steeply to the main basin floor.
The peaks and spikes would be removed by software
processing in a full survey and result from
second time around returns, side echoes from
steep slopes, and from an area of boat moorings.
The plot contains over 200,000 data points.
History and Sediments
The three hundred million-year-old fault was glaciated
by successive ice ages until approximately
12,000 years ago, when the lake we know today
was formed. Meltwater raised the sea level
until the land, relieved of the ice burden,
rose to bring the loch to its present altitude
of 16m. The Loch Ness and Morar Project has
extracted sediment cores from Ness and Morar
in an attempt to shed light upon the post‑glacial
history.
Much of the material remains to be worked but so
far, no evidence of a marine transgression
has been found at Loch Ness. A core from Loch
Morar (9m above sea level) was found by Dr.
H.J.B. Birks (pers. comm.) to contain abundant
cysts of marine algae, which suggests that
the sea could have entered, although the spores
could conceivably have been wind borne.
The deep basin floor of Loch Ness is covered by
black lake sediment, of which a 4m core only
just penetrated to clay, thus suggesting that
the organic sediment is of at least that depth.
This core has yet to be dated, but a 2m core
from the basin floor of Loch Morar has been
estimated by Dr. Birks (pers. comm.), on the
basis of pollen content, to have penetrated
between 5,500 and 6,500 years of post-glacial
sediment. This sheds some light upon rates
of deposition and, incidentally, upon the
likely depth of sediment in Loch Ness. At
the seaward end of Loch Ness the sediment
consists of clays, an interesting feature
of which is that a firm yellow‑brown
layer approximately one metre thick overlies
a blue‑grey material which is very fluid.
The organic content of the loch sediments has been
established to increase with depth (Lee and
Collet, 1908). From an analysis of our own
profundal substrate samples (J.D. Hamilton,
pers. comm.), the percentage composition of
the organic matter ranges from 18.3% to 34%,
similar to the Bathymetrical Survey results.
Like this earlier study, it would also appear
that the Urquhart Basin has less organic matter
than the Invermoriston Basin, which may be
due to the Foyers barrier retaining the organic
material within the southern basin.
1988 The Scottish Naturalist p118
2. THERMAL STRUCTURE
Stratification
In a lake, the onset of stratification in summer
consists of the separation of the upper less
dense warm water, known as the epilimnion,
from the deeper cold water, or hypolimnion,
by a region of sharper temperature change,
termed the thermocline. Photosynthesis is
limited to the epilimnion, where depletion
of nutrients cannot be replaced from the hypolimnion
until mixing occurs in winter (Figure
4a, 7K).
In eutrophic (richer) lakes, the decay of organic
matter descending to the hypolimnion deoxygenates
it, to the detriment of deepwater life. In
deep oligotrophic (poorer) lakes, such as
Loch Ness, these profound effects do not follow,
since nutrients are already low in the epilimnion
and there is no deoxygenation of the vast
hypolimnion, which remains over 80% oxygen
saturated. The oligotrophic nature of Loch
Ness therefore, offers the compensation of
stability in exchange for low productivity.
It is spared the seasonal booms and crashes
of more productive waters, and a variety of
life extends to its deepest regions.
The great body of water does not fall below a uniform
5C in winter, and consequently there
is no inverse stratification and no freezing.
Indeed the influence of the loch's stability
extends to the area around the shoreline,
where the release of heat ensures that snow
seldom lies for long. On 10th January 1987,
for example, the air temperature was 4.4C
after a very cold spell of overnight temperatures
down to -14C. The water temperature was 6.4C
to a depth of at least 50m. Of course the
loch is still relatively cold from a biological
point of view and many of its inhabitants
are relicts from glacial times.
Physical Studies
The biology of Loch Ness may have been neglected
until recently, but physical studies have
been particularly rewarding. The steepness
of the 'walls', length, and regularity of
the basin, together with the loch's orientation
N.E-S.W. in line with the prevailing winds,
all tend to simplify and amplify thermal effects.
The absence of salinity complications has
also facilitated observations in support of
hypotheses afterwards extended to oceanography.
It was during early investigations
at Loch Ness that the first internal temperature
seiches were discovered (Watson, 1904; Wedderburn,
1907). These arise when wind, blowing warm
surface water to the leeward end, tilts the isotherms
lengthwise in this direction. The cessation
of wind stress permits recovery of the isotherms
but, because of momentum, they overshoot and
oscillate for some days (Figures 4a,
4b,
17K and 6a,
6b. 6c,
6d,
approx 12K each)). L.H. Mortimer (1955) of
the Freshwater Biological Association showed
the effect of the earth's rotation on the
currents accompanying seiches, which leads
to a deflection to the right and hence a cross-loch
tilting of the isotherms (Figure
8 , 15K). Mortimer's observations
of asymmetry in the seiche led Dr. S.A. Thorpe
of the National Institute of Oceanography
to establish the presence of a 'surge' wave
front characteristic (Thorpe, Hall and Crofts,
1972). Thorpe (1977) has since conducted other
work, including the mechanisms of mixing and
turbulence.
Factors affecting Sonar
The Project considers it important to understand
the nature of thermal characteristics likely
to affect sonar work. For example, sonar beams
propagating horizontally are refracted by
temperature gradients and it is also important
to recognize direct echoes caused by them.
Conversely, we wish to discover the extent
to which thermoclines can themselves be monitored
by sonar, and their relationship to 'scattering
layers'. Lastly, we are seeking to establish
the role played by physical factors in the
distribution of biomass. The thermocline movements
may affect the range of vertical migrations,
and internal seiches can cause horizontal
transport.
Observations will first be made on the build‑up
of stratification, and then the role of sonar
discussed. From 19th June to 7th October 1983
temperature measurements were made from a
mooring to the north of the deep basin (220m)
off our station just south of Achnahannet
(see map in Figure
1a). The readings were too infrequent
and irregular to give a valid continuous record,
and were complicated by our position near
the node of the main longitudinal seiche,
which doubles the frequency, and towards the
northern side of the basin, where we were
subject to the transverse seiche. An example
of daily records (Figure
5a, 7K) reveals large changes in the
depth and degree of stratification. Nevertheless,
the averages of readings per week (Figure
5b, 6K) serve to show progressive deepening
and convergence of the isotherms towards autumn.
Further background is provided by the record from
the fixed station (see map in Figure
1a) employed in 1984. This was moored
towards the south end of the loch, in the
centre of the deep basin, about 4.5km north
of Fort Augustus, thus simplifying interpretation,
and readings were taken at least four times
per day. The result is shown in Figures
6a, 6b,
6c, 6d.
Winds were measured with a simple hand-held device (Ventimeter) and currents,
where recorded, are merely observations of
drag on the temperature probe wire. A detailed
discussion of the record will not be attempted
in this review but the following comments
may be helpful.
The Record
The record begins on 22nd July (Figure 6a,
12K), as N.E. winds mix and deepen the 17C
epilimnion, until 26th July when the wind
changes to S.W. As the water is transported
to the N.E., successive isotherms break surface,
exposing colder layers which will mix into
the warm water to leeward (Mortimer, 1952).
The epilimnion begins to recover as the winds
moderate on 28th July and with calmer weather,
the upper isotherms generally deepen, although
possibly checked by S.W. winds from 31st July
to lst August, until they are forced down
by strong N.E. winds (Figure
6b,
14K) from lst to 3rd August. As a result of
mixing, the epilimnion is now cooler (14C)
and deeper. Stratification becomes marked.
The disturbances generate a seiche in the lower
isotherms with a period of approximately 54
hours characterized by a rise in temperature
of 18 hours and a fall of 36 hours. The wind
cycle appears to have various amplifying and
dampening effects on the seiche until 13th
August (Figure
6c, 11K) when calmer weather accompanies
a loss of regularity and amplitude. From 15th
August a consistent 15C isotherm re‑enters
the record and an epilimnion is ultimately
deepened by N.E. winds, from 23rd to 24th
August, to bring about a situation similar
to that at the beginning (Figure
6d, 10K).
The process continues throughout the summer as
calms produce transitory shallow epilimnia
which are soon mixed in by winds. Mass transports
caused by seiches spread the heat deeper by
turbulence. The most effective mixing is accomplished
by the autumnal equinoctial gales, which since
the loch is no longer gaining heat, are able
to produce an epilimnion of near uniform temperature.
It is from this simple situation that our
description of the sonar contribution begins.
Figure
7 (19K) shows one of the 252 sonar
profiles made with Simrad EY-M (narrow beam
11 degrees) and Skipper 603 (wide beam 33
degrees) echo-sounders off the
Achnahannet station in 1983. The record for
29th September has an epilimnion at an almost
uniform 11.3C with the thermocline at 40m.
Two temperature graphs serve to show the depth
and degree of stratification to be related
to the echo trace. A striking illustration
of transverse tilt is provided by Figure
8, (15K), where the isotherms conform
to the tilt registered on the trace, although
it would appear that three layers are involved.
An example of mixing occurs during a short period
of high winds from 2nd to 4th August (Figure
9a, 59K). It is not clear whether
a seiche was in progress (although the trace
shows tilts suggestive of currents), but the
temperature graphs before and after the episode
show a cooling and deepening of the epilimnion.
Prior to the autumn situation the gradients
are complex, and another example (Figure
9b, 67K) shows the destruction, or
at least removal, of a secondary thermocline
at about 10m by S.W. winds from 28th to 30th
August.
During the fixed station experiment of 1984, a
Lowrance Eagle Mach 2 was run every hour for
at least five minutes from 31st July to 24th
August and in addition, almost always when
temperature probes were being taken. In
Figures 6e (28K) and
6f (31K) examples of the
records are attached beneath their relevant
positions and often show the thermocline.
From the foregoing it might seem that sonar
is quite reliable in resolving thermal structures
but a considerable degree of caution is required
in use and interpretation.
Possible Errors
Monitoring of open water 'scattering' layers as
early as 1982 had highlighted the problems
of 'second time around' returns, which can
confuse or obscure the record. With range
set to less than water depth, echoes will
return from the lake bed and be received during
the 'listening' phase of subsequent pulses.
If the set range is sufficient to include
the lake bed, second time around echoes can
still be received, having been inter‑reflected
between the bed and water surface. They appear
on the chart as diffuse layers, and are accentuated
by the high gain settings necessary for thermocline
detection. These returns are indicated on
some of the charts, and in practice limit
the choices of effective range settings. Such
returns can be identified since they change
range with alterations in pulse repetition
frequency, i.e. range settings.
Another cause for confusion lies in the frequent
association of biological 'scatterers', such
as fish and plankton (Schroder, 1962), with
the thermocline, at least by day. Detritus
has also been suggested, but we have never
found detectable traces in samples.
In Figure
10 (31K), a comparison of simultaneous
runs with wide and narrow beam sounders clearly
shows the contribution of fish to the wider
beamed sounder's return. In Figures
6e and 6f,
during the fixed station work, many of the
daytime returns from depths of around 30m
are strengthened by the layer of fish. For
further discussion see sections on plankton
and fish.
1988 The Scottish Naturalist
p126
The records sometimes show inconsistencies between
the degree of stratification and the strength
of echo trace. It is particularly noticeable
that, on the narrow‑beam Simrad EY‑M,
thermal gradient traces often appear patchy.
An examination of Figure
8, (15K) for the southern profile
shows discontinuities and traces to be reasonably
consistent, but to the north, strong traces
register from lesser gradients. This may be
partly accounted for due to the thermocline
being divided into 'layers' of weak gradient,
several metres thick, divided by much thinner
'sheets' with steep gradients (Woods, 1968;
Simpson, 1970). Our one‑metre temperature
sampling interval would not resolve microstructure
adequately. We believe, however, that the
explanation lies in turbulence, to which echo‑sounders
are particularly sensitive, as shown by traces
throughout the paper recording the raising
and lowering of samplers and illustrated vividly
in Figures
9a and
9b showing mixing. In the above
example the transverse tilts, by definition,
imply strong currents moving in opposite directions.
Echoes are often pronounced near the side
walls where greater turbulence is to be expected.
Shear
Shear is actually associated with most thermoclines
and waves within them sporadically produce
inversions known as Kelvin-Helmholtz billows
(Woods, 1968; Thorpe, 1974). We believe this
turbulence to be the main factor in regulating
the strength of echo returns, possibly by
creating multiple interfaces of small size
but high gradient. In support of this, attention
is drawn to the 1984 fixed station record
in Figure
6f (31K) for the period 3rd to 7th
August, which includes the sharpest thermocline
observed. The records for the night of 3rd
August show the gradient strongly after the
fish have left it and while the isotherms
are rising. However, by the time of the temperature
profile at 10.00 hrs the interface echo was
weak or undetectable despite the intensity
of stratification. It is at this point that
the isotherms are at rest and the currents
therefore at their slackest. A conclusive
thermocline trace does not re‑enter
the record until 5th August at 22.00 hrs,
when the isotherms are once again in significant
motion.
The foregoing conclusions seem particularly relevant
to our observation in October 1985 of a surge
front wave of the type described by Thorpe.
These waves (Figure
11a, 12K) lead the seiche and have
a length of approximately one kilometre and
a speed, depending on amplitude, of approximately
1.5 km/hr. Figures
11b, 11c,
11d,
11e,
11f,
show a continuous echo trace from 18.55 hrs
on 11th October to 14.44hrs on 12th
October, superimposed with some relevant temperature
graphs and current observations, all taken
from a moored position close to that of the
1984 fixed station. S.W. winds on the previous
night had reached hurricane strength over
parts of Scotland but had calmed to a gentle
breeze by the time we took station. The record
commences (Figure
11b, 115K) with a shallow 12-15 metre
epilimnion of a uniform 10C. At 23.15 hrs
(Figure
11c, 76K) the thermocline dipped sharply
and a spectacular wave train of decreasing
regularity was observed. Temperature probes
(Figure
11d, 93K) show a rapid lowering of
the isotherms with discontinuities corresponding
to the echo trace, and at 01.15 hrs an increasing
subsurface current to the S.W. set against
the surface drift. By 02.28 hrs the surface
water was also moving against the wind and
by 08.00 hrs (Figure
11e, 75K) was rapid enough to cause
difficulty with temperature probes and plankton
sampling, because the wires assumed unacceptable
angles. At 13.07 hrs (Figure
11f,
86K) the current was easing, and at 14.30
hrs was slack. By this time the S.W. wind
had freshened and the surface drift had reverted
to N.E.
Obviously the entire event was attended by a high
degree of shear as the epilimnion slid beneath
our station and it will be noted that the
surge front wave crest registers particularly
strongly. We speculate that the crest comprises
turbulence similar to that observed by Woods
(1968) on thermocline sheet waves, although
of a greater amplitude. The two initial waves
are of 23m and 45m amplitude. The trace from
06.30 hrs to 08.15 hrs (Figure
11e),
when the strongest currents were observed,
is interpreted as consisting of Kelvin-Helmholtz
billows (Thorpe, 1988). The cause of the near
surface turbulent effects seen from about
08.15 hrs is less obvious but coincides with
the influx of slightly warmer surface water
and is probably also associated with shear.
The plankton hauls (Figure
11g,
10K) show a mass influx at 02.08 hrs, and
it is evident that fish are also being transported,
sometimes in very localized concentrations
(see sections on zooplankton and fish).
Conclusion
In conclusion, it may be said that echo‑sounders
have considerable value in the detection of
general thermal conditions, but do so by registering
degrees of turbulence rather than gradient,
even though the two are often associated.
Future experiments will include induced artificial
turbulence, which may then propagate across
potentially unstable structures, thus revealing
their positions (see Figure
12, 12K).
3. BIOLOGY
A measure of the neglect shown towards Loch Ness
may be judged from the fact that the first
basic observations to be made there since
the Bathymetrical Survey in the early years
of this century were made over fifty years
later, by teams investigating the 'Monster'
controversy. Student expeditions from Birmingham
and Cambridge Universities made plankton hauls,
while Mackal and Love (1970) of the Loch Ness
Investigation Bureau took water samples (Figure
13a).
The situation changed, however, when from 1977
to 1980 a comparative study of five major
lochs, including Ness and Morar, was undertaken
by the Institute of Terrestrial Ecology. Wide‑ranging
papers were edited by Maitland (1981) in the
book The Ecology of Scotland's Largest
Lochs. The Project's general biological
objectives have therefore concentrated upon
areas not covered by the comparative study,
such as plankton and fish migrations. A special
study has been made of the abyssal fauna.
Phytoplankton
Primary productivity is low. The high latitude
and frequent cloud reduce sunlight to a short
growing season. Photosynthesis is further
limited, by suspended peat, to a shallow photic
zone. Rooted plants are restricted to a depth
of about 6m around the shoreline. The hard
rocks of a steep catchment yield few nutrients
to fast flowing rivers and streams entering
the loch. Acidity will tend to slow bacterial
decay of organic particles and hence the release
of their nutrients (Figure
13a, 12K).
Thus the phytoplankton crop is low, and was found
by the Cambridge expedition to have a diatom
population of 200/1 as compared to 5,000/1
for Lake Windermere (Baker, 1962). Bailey-Watts
and Duncan (1981) found the plankton in Loch
Ness to be dominated by chrysoflagellates,
with a total cell count of 568/1 at the time
of the chlorophyll 'a' peak in August. Some
of the species are shown in Figure
13b (19K).
Zooplankton
The cladoceran grazers of the phytoplankton consist
of Diaphanosoma brachyurum, Holopedium
gibberum, Daphnia hyalina and Bosmina
coregoni. The larger predators are Polyphemus
pediculus, Bythotrephes longimanus and
Leptodora kindti. Copepods are represented
by Diaptomus gracilis, the most numerous
species, and Cyclops strenuus abyssorum.
See Figure
13c (16K).
Zooplankton of most open waters appear adapted
to avoid predation by transparency, and at
Lochs Ness and Morar are also adapted to low
productivity, by remaining relatively small
and by producing fewer but larger eggs than
their counterparts in more productive waters.
Food seems to be stored in some copepod specimens
as oil globules. Peak numbers have been found
in October (Maitland, 1981: 144). Project
work was conducted in collaboration with Dr.
A. Duncan of Royal Holloway College in September
1983, with the aim of determining vertical
and horizontal distribution. A Clarke‑Bumpus
collector was used to take horizontal hauls,
which have yet to be worked, and a 31‑litre
Patalas sampler was used for the vertical
work.
The vertical hauls are shown in Figures
14a
(49K) and 14b
(40K) with a summary in
14c (16K). From 12.00 hrs on 12th
September 1983 to 05.00 hrs the following
morning, five hauls were made, in conjunction
with temperature profiles and echo‑soundings,
from the mooring off the Achnahannet station.
The results show a clear concentration of
plankton at the thermocline at 13.00 hrs which
involved most species. Thereafter, a migration
takes place towards the surface, particularly
in the case of Cyclops and Diaptomus. Total
numbers increase towards midnight, possibly
due to horizontal transport in response to
the rising of the isotherms. Some of the peak
densities are noted and have no convincing
connection to the echo traces, which, although
complicated by second time around returns
and reflections from the mooring, appear to
bear more relationship to the temperature
profile.
There has been
considerable research on acoustic scattering
by marine zooplankton. Theoretically even
a 200kHz echo‑sounder of high sensitivity
could detect layers of organisms as small
as one millimetre, and euphausiids of 15‑22
mm have been assessed by multifrequency methods
(Greenlaw, 1979). In fresh water, where plankton
sizes are generally much smaller, convincing
records have been made at 200kHz of Chaoborus
larvae in the absence of thermoclines
(Northcote, 1964). However, these insect members
of the zooplankton have a length of 9mm-12mm
together with paired air sacs at either end,
contributing to much higher scattering strength
than from the more typical cladocera and copepods.
Chaoborus is absent from Loch Ness,
and the largest of the more abundant herbivores
measure up to 2mm.
Schroder (1962) reported the detection of one millimetre
freshwater plankton at densities of as little
as 2/litre, although the sonar frequency was
not stated. Examination of the traces suggests
that plankton horizons presented cannot be
distinguished from the thermocline records.
Indeed, the prime conclusion of the paper
is that the "zooplankton in Lake Constance is mostly found by the echo‑sounder
in layers with sharp gradients (thermoclines)
where it is also to be found during vertical
migration". At our frequency of 50kHz
(33 degree beam) no correlations
were evident at concentrations of 10/litre.The frequent concentration of zooplankton
in thermoclines must also inevitably associate
them with the temporal and localized shear
to which echo-sounders are so sensitive. It
would seem difficult to design thresholds
for echo integration of zooplankton volume
scattering where thermal gradients exist.
An example of horizontal transport appears to occur
in the strong currents noted during the surge
shown in
Figure 11d (93K). Four sets of net
samples were made, all but the first to a
depth of 36m in 6m stages. A closing net of
28cm diameter was used. The results indicate
a mass influx of plankton as the currents
strengthened.
To conclude, we have observed zooplankton in Loch
Ness concentrated at the thermocline by day
and exhibiting a vertical migration at night.
It may be of interest that this migration
has not been observed in Loch Morar, where
the water is much clearer but where there
are far fewer midwater fish (Figure
14c, 16K); here the plankton remain
in the top 15m of water by day. In Loch Ness
considerable horizontal transport of plankton
occurs, particularly during a seiche. We are
not convinced that, at the concentrations
observed, our echo‑sounders have yet
made a contribution to recording zooplankton
(50kHz‑200kHz) and draw attention to
possible confusion owing to the presence of
turbulence. Layers of small fish may also
give misleading (diffuse) returns, particularly
at depth and on wide‑beamed sounders.
Diagnosis is possible if a vertical migration
takes place (see Figure
20a, 13K).
Littoral and Sublittoral Benthos
The steep and stony shores of Loch Ness provide
a restricted habitat with considerable exposure
to wave action (Figure15a,
(7K). The fauna is therefore similar in some
respects to that of fast‑flowing streams,
in that the organisms must retain their positions
among stones (Figure
15b, 11K), rather than on the silts,
muds and plant life of more lentic waters.
A comprehensive survey
of the animals dwelling within the first 50cm
of water, conducted by the Institute of Terrestrial
Ecology (Maitland, 1981) confirms that the
community is dominated by insect nymphs. Stonefly
(Plecoptera) make up 30% and mayfly (Ephemeroptera)
18% of the fauna, some of which are adapted
to retain position by the possession of grasping
claws or present a low profile to the water
by a dorso‑ventral flattening of their bodies. A common mayfly, Ameletus
inopinatus, is generally confined to streams
above 300m or to lochs much further north,
so its presence here suggests a preference
for the lower temperatures found in deep,
windy lakes, even at the surface, owing to
regulating effects of mixing.
Other benthos include triclads, various nematode
and oligochaete worms, the gastropod Lymnaea
peregra, the crustaceans Asellus spp.
(found in more sheltered areas) and ostracods,
water mites, and caddis (Trichoptera) and
chironomid larvae (Figure
15c, 25k). Typical Project hauls at
Achnahannet and the Horseshoe Scree ‑
additional to Maitland's list ‑ are
included in Figure 15d
(10K).
The steeply sloping walls of Loch Ness provide
a narrow and ill‑defined sublittoral
zone, but hauls from 20m‑30m depths
at Urquhart Bay as well as Borlum Bay, in
sandy sediments, have revealed the organisms
listed in Figure 15e (10K). These littoral
and sublittoral communities, together with
terrestrial insects falling into the water,
provide the food for the inshore fish populations.
Profundal Fauna
In contrast to the turbulence and variety of physical
conditions among the stones of the shoreline,
the fine and relatively rich silts of the
abyssal regions offer remarkable stability.
In an environment of great hydrostatic pressure,
constant darkness, and a scarcely changing
low temperature of 5.6C, high oxygen levels
(over 80% saturation), permit surprising variety
in the profundal community of the 200m deep
basin floors.
The first samples were taken during the Bathymetrical
Survey (Murray, 1904: 442). It would appear
however, that no further collections have
been made until the present work. For a description
of the sediments and a literature review of
the benthos see Maitland (1981: 205). The
Project has made a particular study of the
profundal fauna at both Ness and Morar and
a few comments are appropriate here.
A variety of qualitative
and quantitative collection techniques have
been used, including dredges, grabs and a
static 'colonization experiment'. At Loch
Ness, over thirty species have been recorded,
some of which are doubtless casual occurrences,
with an average density of 295 individuals
per sq m. Recognized characteristic
profundal fauna is present, such as oligochaete
worms, chironomid larvae (non‑biting
midges) and Pisidium spp. (pea mussels)
but, numerically, ostracods predominate, comprising
62.6% of the community. The dominant bivalve
mollusc, Pisidium conventus, is more
normally found in arctic streams, but at our
latitudes is confined to the cold water of
deep lakes or to high altitudes. It is considered
to be an ice-age relict species in Loch Lomond
(Hunter and Slack, 1958). The dominant chironomid,
Sergentia coracina has been similarly
described in southern Sweden (Brundin, 1949).
The community also has its predators, such
as chironomids of Procladius spp. and
the large copepod Acanthocyclops viridis,
which are more cosmopolitan in distribution
although apparently absent from the littoral
zone of Loch Ness.
Some interesting occasional records include caseless
caddis and the flatworm Phagocata woodworthi,
which has been recently discovered in
the littoral near a sewage outfall (Reynoldson,
Smith and Maitland, 1981). This North American
triclad is speculated to have been introduced
on equipment imported in the search for the
Loch Ness Monster. Another native of North
America, the amphipod Crangonyx pseudogracilis,
has been spreading northwards since its
discovery in the London area (Crawford, 1937),
and its occurrence during the present work
is the first record for a Scottish loch. It
could well have been responsible for small
fast moving images first seen on underwater
television in 1981. It may be significant
that Crangonyx was captured only in
the passive colonization substrate. Fish,
again first noted during the television work
of 1981, have now been netted and identified
as Charr Salvelinus alpinus. Figure
16a (24K) shows some of the members
of the abyssal fauna.
The role of the echo‑sounder during this
work was limited but aside from enabling the
quick location of the depths to be sampled,
sonar can often aid the control of sampling
equipment. In particular, it was noted that
the Ekman grab penetrated too deeply to enclose
the vital top 5cm of sediment. With the echo‑sounder,
sampling was quicker and more effective since
the sampler could be allowed to free fall
to within a few metres of the bed and then
be gently lowered. Figure
16b (10K), shows a grab being deployed,
and also a method for the recovery of long
term experiments, in this case our colonization
substrate sampler, by the location and grappling
of a buoyant rope laid horizontally and with
the end weighted.
In general, it seems that the depths offer a refuge,
not only for ice‑age relicts but also
for a variety of widely occurring 'pond life'
unsuited to the turbulence and predation pressures
of the shoreline. It will now be necessary
to examine the rocky side walls to define
the limits of the two communities and to see
if there is also a third.
Fish
As the ice retreated, the loch was colonized from
the sea by coldwater salmonids, which entered
fresh water only to spawn. Thus the Salmon
Salmo salar still migrates via the
River Ness, as does the Sea Trout Salmo
trutta. However, with the passage of time
and increasing sea temperatures another variety
of Salmo trutta, the Brown Trout,
and the Charr Salvelinus alpinus now
spend their whole life cycles in the loch.
It seems possible that new populations of
Charr may have entered during the period of
the Loch Lomond re-advance (Greer, pers. comm.).
Other species gaining access
by sea migrations are the Three‑spined
Stickleback Gasterosteus aculeatus, the
Brook Lamprey Lampetra planeri and
the Eel Anguilla anguilla, which
has an opposite life cycle to the salmonids
in that it matures in the loch after having
been spawned in the Sargasso Sea.
The exclusively freshwater 'coarse fish', which
have been spreading northwards since the ice
age (Maitland, 1977), are represented only
by the Pike Esox lucius and the Minnow
Phoxinus phoxinus. The Minnow
was not recorded by Maitland (1981); it was
first observed by the Project in 1985 at Bona
Narrows just downstream of the loch proper,
but in June 1987 it was also recorded in sheltered
water at Fort Augustus.
In collaboration with members of the Ness Fisheries
Board, together with Dr. A. Duncan and Mr.
R. Greer, various netting experiments have
been carried out. Underwater television (UW/TV)
has so far made contributions in the littoral
and abyssal zones, while sonar has been most
extensively used in the pelagic. Much work
remains to be done on the ageing and electrophoresis
of the specimens.
Littoral Zone
The littoral is dominated
by brown trout to a depth of about 20m, below
which charr prevail. The really shallow and
sheltered water harbours minnows, three‑spined
sticklebacks and salmonid parr. Eels live
on the loch bed and range widely along the
shoreline, being most concentrated off river
mouths. Work on the growth rate of the Loch
Ness eels has been conducted by Mackal and
Frake (Mackal 1976: 319‑330).
Benthos is doubtless an important food source and
many of the brown trout seem particularly
specialized, e.g. a 21.7cm fish contained
143 snails (Lymnaea peregra). In summer,
however, a substantial proportion of the trout
diet consists of a terrestrial input of adult
insects. Figure
17a (27K) presents some relevant information
on the fish of the littoral. Figure
17b (16K) shows underwater television
pictures of trout and eels in Urquhart Bay,
concentrated in the rich organic leaf deposits
at 30m off the mouth of the River Coiltie
in early June 1987. As summer progresses. a considerable amount of gas generates
in this material and also in other parts of
the bay, possibly because of higher water
temperatures. In October 1987 only an occasional
Eel was observed here. Gas has not been detected
in the fine deepwater sediments.
It is along the shoreline that the migrating salmon
pass before spawning in rivers, and attention
is drawn to the fact that salmon, as the largest
fish present, owe very little to the loch's
rather poor food chain. After only a few years
as small parr they will then mature at sea,
and return at weights up to 20kg. There is
no fish counter on the River Ness, but two
counters installed on the Rivers Garry and
Moriston showed less than 900 fish in a peak
year of 1975 (Maitland, 1981: 243). Although
this does not include the rivers in Urquhart
Bay, a comparison may be made with a maximum
count of just over 3,000 fish entering Loch
Morar in 1966. The picture is rather sad when
one recalls the comments of Captain Burt at
Inverness in 1758, that the price of salmon
was a penny a pound and that "the merest
servants who are not at board wages will not
make a meal upon salmon if they can get anything
else to eat" (Mills, 1980).
Profundal Zone
We have yet to
observe fish by underwater television on the
loch 'walls' beneath about 150m. The fish
of the profundal are not amenable to detection
by sonar because of attenuation and widening
beam angle, so the information presented
results from UW/TV (Figure
18a, 7K) and netting experiments at
180m-220m depth on the deep basin floors(Figure
18b, 7K). Future experiments will
include lowering deep‑towed narrow‑angle
transducers to assess this region.
The presence of deepwater fish was confirmed in
1981 through UW/TV, and in 1982 the first
three specimens of charr were netted from
220m. It is always something of a problem
to be certain that fish are really caught
at depth and not during the long setting and
retrieval process, but of the twelve charr
caught at this depth, seven contained profundal
fauna including Pisidium conventus. Some
fish believed to have been caught at 200m
had surface zooplankton in the lower gut,
thus suggesting that a relatively fast descent
takes place (Figure
18c,
15K). Versatility is thought to enable Charr
to inhabit relatively sterile lakes and to
avoid direct competition with the less adaptable
brown trout.
On the other hand, it has been shown that there
are two distinct races of charr, a pelagic
and benthic form, in Loch Rannoch (Walker,
Greer and Gardner, 1988). Electrophoresis
has shown genetic differences between the
stocks and initial results with our Loch Ness
specimens also appear to show two varieties, although the visible differences, larger
eyes for example, do not appear obvious at
this stage.
Pelagic Zone
The
offshore waters to 30m are dominated by charr,
which feed upon the larger zooplankton such
as Daphnia, Leptodora and Bythotrephes.
As part of the fixed station work of 1984,
depth‑marked nets of various meshes
were suspended beneath the raft to a depth
of over 30m, aligned N.E.-S.W. The results
in Figure
19a (17K), show a poor catching frequency
between 8th and 21st August with charr taken
at approximately 10m by night, when it is
also evident that trout extend over the surface.
In addition to zooplankton, the trout contained
a proportion of winged insects (Figure
19b, 26K). The only fish caught by
day were two trout at approximately 30m; one
of the trout contained two charr and may have
been a "ferox".
Although in general the catches tend to support
the presence of vertical migration, it should
be borne in mind that echo‑sounding
shows individual fish within the net depth
by day, and it seems likely that avoidance
was significant. No monofilament net was used.
The catch depths could well be a result of
the depths at which the net was visible to
given species, or of their periods of maximum
activity. Nevertheless it seems from this
result and from much longer nets set on the
surface that charr, at least specimens of
over 20cm, rarely come within three metres
of the surface even at night.
Echo‑sounding with a variety of instruments
reveals the basic pattern of summer activity.
Figure
20a
(13K), shows that by day, a scattered
population of individual fish range down to
the thermocline. Rough calibration suggests
these fish to be over 20cm in length. In the
region of the thermocline a well defined layer
of fish at greater density requires a narrow‑beam
sounder to resolve individuals (Figure
20b, 16K), which may otherwise blur
into the thermocline traces. The calibration
suggests that this layer includes fish which
are very much smaller than the scattered population
and which would not have been caught by our
net meshes. These are mainly charr, including
the 0+ age group (confirmed by trawling during
1988), but we have also made some interesting
incidental catches of trout and salmon parr
in open water. At dusk, this layer makes a
distinct vertical migration and by midnight
it is within six metres of the surface. It
seems that little dispersion takes place and
at dawn the layer sinks. Again, as by day,
a scattered population of individuals is detected
at depths to 30m unrelated to the layer.
Figure
20d (42K) details the limits of the
vertical migration as observed from the 1984
fixed station between 31st July and 24th August.
Unfortunately the above netting programme
obscures parts of the record. Throughout the
period the layer descended to a maximum of
35m, usually sufficing to bring it within
a temperature of 10‑11oC
and always to under 12oC.
It descended to at least 30m, however, even
though it may have reached the 12C
isotherm at a lesser depth. Some degree of
temperature preference would seem to be shown
by the deeper (35m) daytime depth on 3rd August
when the isotherms are depressed. The fish
do not necessarily seek out or remain in thermoclines
(see 4th August) but there is more
chance of any preferred temperatures being
'compressed' into this area. In the generally
cooler epilimnion temperatures (and overcast
conditions) during the summer of 1988, the
layer's daytime depth could be as little as
18m (at a temperature of 12.5C), well
above the thermocline.
Much work remains to be done at other times of
year. In the meantime, isolated observations
suggest that the migrating layer becomes established
in late May, although its daytime depth is
only 24m. An interesting question to determine
is whether this is due to lesser light levels
or to cooler epilimnion temperatures. The
migrating layer seems to disperse around October,
when scattered fish may be detected to depths
of 50m or more. The winter mid‑water
fish population is reduced.
Regarding the horizontal transport of fish (Figure
20c,
27K), attention is also drawn to the surge
record in Figure
11c
(76k). Since the soundings are from a
moored position, the speeds at which a fish
at a given depth crosses the beam will shorten
or lengthen its characteristic arched signature.
Fish at greater depths will obviously spend
longer in the beam because of its width.
It is noticeable that the influx of fish exhibits
compressed arches, particularly when the strong
currents were noted, in contrast to the generally
long arches shown beforehand. This is suggestive
that the movement is a passive one due to
the currents, rather than active swimming.
Harden‑Jones (1969: 16) suggests that
in the absence of visual cues, fish are unable
to detect laminar flow, but this could change
in the presence of a discontinuity in the
current, such as shear
A further point is that even a simple echo‑sounder
can resolve horizontal movement in one plane,
without affecting vertical accuracy, by giving
a small tilt to the transducer. Thus, fish
traces will show asymmetrical uprange and
downrange components, depending upon their
direction of movement along the beam axis.
Even a vertical transducer is likely to have
some bias to the beam. This is very noticeable
in the surge tracings where fish show a distinct
downrange trend. On this occasion the bias
of the transducer was not known, but it seems
likely that the traces show a movement in
conformity with the current. It will also be observed that fish are
concentrated behind internal wave crests.
Fixed station records in Figure
20c
(27K) also show a bias but with periodic
changes of direction in parallel with seiche-induced
currents.
During Operation Deepscan in 1987 an attempt was
made to map fish distribution after the break
up of the scattering layer. At 12.00 noon
on 7th October, seventeen craft
(equipped with Lowrance X-16 sounders) made
simultaneous profiles, sectioning the length
of the loch. Each then made a run in mid-loch
to the next station, thus yielding a continuous
17.5km record within a 15minute time span.
It had been hoped to chart the thermocline
at the same time but despite maximum sensitivity
being applied, this was not detected. We surmise
that this was due to the calm weather reducing
the movement of the isotherms and resulting
shear. Temperature probes over the period;
show an epilimnion of a uniform 11.5C
to 30m.
Figure
21 (12K) shows the results of visual
fish counts and although no replicates were
made, it appears that some interesting variations
in density were present. There are more midwater
fish well offshore and numbers increase towards
the S.W. end of the loch. Further research
seems worthwhile on the role played by physical
factors in fish distribution. At the same
time, it would be valuable to discover the
periods of peak activity of the fish forming
the layers, possibly by lowering transducers
from fixed stations to keep range and angle
constant.
It would be desirable to adopt more sophisticated
acoustic estimation techniques but our observations
highlight some possible problems (Figure
22,
33K). With scattered fish, it is customary
to adopt an echo-counting system, which becomes
less useful as density increases, as in the
scattering layer, because of overlapping signals.
The method used for higher densities, echo-integration,
is liable to suffer inaccuracy because of
these fish being found in the region of the
thermocline with its associated echoes. By
night, fish which have risen close to the
surface, could be scattered by the approach
of the survey vessel.
In August 1988 the scattering layer lay at 18-20ms,
well above the thermocline and echo-counting
with relatively few (30%) overlaps was possible.
A Simrad Hydro Acoustic Data Acquisition System
(HADAS) was interfaced to the output of an
EY200 (49kHz) Scientific Sounder. The subsequent
software analysis has measured area densities
of up to 1,000 fish/hectare within the layer.
Part B: THE MYSTERY
1 THE CONTROVERSY
Introduction
The work of the
Loch Ness and Morar Project during the 1980s
reflects evolution not only of method but
also of attitude to the Loch Ness controversy.
The history of organized searches during the
1960s and 1970s has been described by Fitter
(1988); see also Dinsdale (1961), Mackal (1976)
and Rines et al. (1976). Although successive
investigators remained impressed by a portion
of eyewitness testimony, it must be understood
that, by the end of that period, most experimental
evidence had been subject to review and to
a degree of rejection. Some of these alternative
interpretations were ultimately to become
available in book form (Binns, 1983; Campbell,
1986; see also Razdan and Kielar, 1984).
The evolution of method itself bears tacit witness
to a revision of expectations. By the end
of the sixties, intensive surface surveillance
gave way to underwater work, after having
failed to reproduce any of the highly varied
'classic monster' photographs (Mackal, 1976:
122). In turn, for all their early promise,
underwater camera vigils at Lochs Ness and
Morar had been discontinued by the late seventies.
The realization that passive underwater cameras
were not going to provide an easy positive,
let along negative, answer could only prelude
a more active approach and a long period of
verification for the single category of experimental
evidence, which seemed repeatable and for
which no satisfactory explanations had been
put forward.
Sonar,
although a long range tool, lacks definition,
and its prime role lies in demonstrating the
presence of something of interest in the first
place. In turning to sonar and examination
of its previous contributions, the Project
acknowledged that such a demonstration was
after all necessary. The motives for experiment
have also changed. Fairness to the eyewitnesses
may justify research, but other evidence hardly
merited a position of advocacy. At Loch Morar,
Project work had already diversified to contain
a major component of general limnology, which
in a revisionist climate, did much to justify
continued organized expeditions. Effort shifted
in 1980 from underwater cameras in the clear
waters of Loch Morar to sonar in the uniform
basin of Loch Ness. The objective was seen
in terms of exploring a neglected habitat,
the presence of which gave grounds for controversy
but in which that controversy played only
a part. Thus the problem is being tackled
through active examination of the environment, rather than
by all or nothing ambuscade.
Work in the 1980s is not a quest for the dragon
of popular expectation. The media‑christened
'Monster', by definition imaginary and by
connotation prehistoric, has an existence
in the realms of entertainment copy, quite
independent of research findings (Meredith,
1977: 156). Admittedly some 'Gothic revivalist'
expectations emerged during the seventies,
as described in a perceptive apologia by Bauer
(1986). The concept, however, had received
little fuel from previous fieldwork, such
as Baker's (1962) dismissal of the huge multi‑humped
stereotype as boat wake effects, or from Mackal's
(1976) analysis of possible candidates in
the light of ten years of research by the
Loch Ness Investigation Bureau. Examination
of the controversy through the environment
reveals some paradoxes between ecology, evidence,
and the choice of operational method.
Biological Considerations
Habitats
Despite low productivities
it is clear that Loch Ness could support resident
or migratory fish predators. The months which
returning Salmon spend in fresh water before
spawning represent a bonus to the food chain
which would once have been very substantial,
and even now provides sufficient incentive
for seals to enter (Williamson, 1987). In
documenting a Common Seal Phoca vitulina
present from 16th November 1984 to 11th
June 1985, Dr. Williamson points out, incidentally,
that it was readily identified by about thirty
people and that no increase in Monster sightings
was noted. If migratory fish are considered
as a food source, one might expect predator
activity in the littoral zone. Mackal's analysis
(1976: 85 and 346) concludes that 52.7% of
sightings occurred in bays around river mouths
but recognizes that these may result from
human activity being greater at such places.
See also sections on Submerged and Floating
Logs.
It was nevertheless
on sound biological principles that the littoral
was chosen for the underwater vigils during
the 1970s, at Loch Ness by time lapse photography
(Rines et al, 1976) and at Loch Morar
by silhouette underwater television (Shine,
1976). Results at Loch Morar were negative
and at Loch Ness were to prove controversial.
No such work has been continued.
The charr of the open waters also represent a large
biomass, since they enter the food chain at
a lower level than other fish of their size,
feeding directly upon zooplankton without
the intermediate link provided by the 'foragers'.
They are already subject to predation by the ferox trout.
If larger predators were to utilize this source,
they should be sonar detectable. By day (in
summer) a depth of 30m may be expected, with
a possible upward migration at night.
The paradox of the loch's oligotrophy actually
extending the range of usable habitat has
been discussed. The community of the profundal
zone has provided unexpected variety and sufficient
concentration to support its own fish population.
This region is very large, because of the
mean depth, and although it would be premature
to propose the basin floors as habitats for
anything unexpectedly large, perhaps they
should not be overlooked either. Benthic creatures
would be difficult to detect by sonar unless
excursions into the water column occur. It
may be worth pointing out that benthic fish
are likely to exhibit peculiar form and behaviour
if they do occasionally surface.
At present, the least hospitable regions would
appear to be the side walls, although these
are difficult to assess by sonar, and the
vast water column between the profundal and
pelagic zones. Yet it was here that the Loch
Ness Investigation Bureau were to achieve
their sonar results
Theoretical relationships between biomass and potential
Monster production have been explored by Sheldon
and Kerr (1972), calculated from Ryder's (1964,
1965) Morphoedaphic Index (total dissolved
solids/mean depth); also by Scheider and Wallis
(1973) using a size‑density relationship,
and the method of least squares, to fit a
power function to data from a marine area
of low productivity theoretically similar
to Loch Ness.
Both authors estimated a similar biomass, for the
terminal predator feeding on fish, as 15,675kg
and 15,725kg respectively. The true figure,
however, is likely to be below that of Sheldon
and Kerr's, because they used data from the
slightly richer Loch Lomond, but neither
figure takes into account migratory fish,
which allows greater biomass. Terrestrial
inputs, e.g. allochthonous leaf material and
winged insects, should not be underestimated
either.
The Candidates
There is considerable speculation concerning
the identity of the Loch Ness Monster; put
another way, this means seeking an explanation,
perhaps many explanations, of the sightings
record. Assuming for a moment that there may
be an unusual single animate explanation,
and confining discussion to vertebrates, environmental
factors are as follows.
Low water temperatures are evidenced by the presence
of cold stenothermic and holarctic species.
As shown in Figure
5b, epilimnion temperatures above
12C are present for only four months of the year. Although the winter temperature
of about 5.6C is actually higher than in shallower
lakes further south, reptiles would seem to
be eliminated. Mammals (e.g. pinnipeds)
would presumably have already drawn attention
to themselves by their more frequent need
to breathe.
Freshwater invertebrates spread rapidly to favourable
habitats, but it has been emphasized that
the connection to the sea has been virtually
the only avenue for colonization by fish.
Salmon, Sea Trout, Lampreys and Eels still
migrate via the River Ness. Ten thousand years
after the final passing of the ice, the European
coarse fish have yet to arrive; indeed the
varieties available in the British Isles are
limited through the inundations of the present
North Sea and English Channel about 7,500
years ago.
The amphibian hypothesis would require a uniquely
large and exclusively aquatic species to spread
rapidly north in the wake of the ice and to
suffer equally rapid extinction elsewhere.
If, as seems reasonable, we look to the sea
for unusual residents or visitors (the amphibia
have no marine representatives) then the fish
would seem to offer the most likely candidates.
Known fish are quite capable of answering the criteria
of size (median length 4.57m: Mackal, 1976:
344), the obviously uncharacteristic nature
of surface appearances, the problems of access
or reproduction, and just about everything
else. Were the eyewitnesses to find vindication
in the discovery (say) of Sturgeon Acipenser
sturio, or indeed any vindication,
most would doubtless be content. Science might
be delighted with the presentation of an outsized
eel (the original local interpretation incidentally),
but perhaps to 'Monster hunters' ten years
ago, this would be "not quite Nessie".
This does not mean that other explanations are
impossible but at least an acknowledgement
of the most plausible candidate from an environmental
point of view, has done something to bridge
the divide between science and 'fringe', which
opened in the 1970's as described so well
by Bauer (1986). The original newspaper report
(Northern Chronicle, 27th August 1930)
described a "fish .... or whatever it
was". If we look for a fish 'on principle'
, we are unlikely to miss anything else, because
we are going to have to look everywhere. Examination
of Loch Ness has proved rewarding in itself.
Physical Factors and Inanimate Explanations
Optical Effects
Before looking for unusual
animate causes for the sighting record, our
observations allow comments upon :some suggested alternatives.
Logs, boats and boat wakes, windrows, otters,
swimming deer, and ducks are all documented
as producing Monster sightings
(Binns, 1983: 165-200). These phenomena are
more noticeable in calm weather, when even
ducks can generate wakes apparently out of
all proportion to their size. On a vast sheet
of featureless water no accurate assessment
can be made of size or speed for an unfamiliar
object, since there is no visual clue to range.
Mackal's (1976) analysis rejects all but 258
out of 3,000 sightings.
Physical effects can further distort perceptions.
The temperature regime is fundamental to mirage
effects (Figure
23a, photo 9K) noted by the Bathymetrical
Survey (Murray and Pullar, 1908a and 1908b),
and the relevance of this to lake monsters
is discussed by Lehn (1979). It was the recognition
of a mirage illusion, which led witness Alex
Campbell to the original withdrawal of the
archetypal plesiosaur sighting (Gould, 1934:
100-113). This sighting has been more influential
than any other in supporting the Jurassic
hypothesis of monster lore.
Professor Lehn
described optical distortions affecting observations
of familiar objects, such as branches, through
direct non‑uniform thermal gradients
in the air overlying cold lakes. The most
usual effect is to extend the image vertically,
but it may also be compressed, appear to move
sinuously and may finally disappear without
a ripple. Optical distortions occur in the
calm weather associated with Monster sightings,
since lake temperatures can then more readily
influence air temperature in the lower strata.
The most pronounced distortions occur
when the observer is close to the water level.
It is suggested that Loch Ness water temperatures
will be significantly lower than the air temperatures
for the first half of the year, thus guaranteeing
strong gradients. This is actually an oversimplification.
For the first quarter of the year (until March-April),
water temperatures are actually above the
mean air temperature, and the Bathymetrical
Survey describes mirages as a winter and spring
phenomenon, attributable to warming of the
lower layer of air (Murray and Pullar, 1908b).
The consequent temperature inversion, although
formed by an opposite process, still has the
effect of distorting images vertically.
Fixed station records from 1984 (Figure
23b, 13K) show the moderating influence
of the loch upon air temperatures, when compared
to the maximum, minimum and 09.00 hrs readings
from St. Benedict's Abbey weather station,
situated on the loch shore 8.6m above the
water level. It will be seen that, even in
summer, the air temperature falls below water
temperature at night. By dawn an inversion
will result, giving rise to 'desert' type
mirage effects. By mid‑morning a direct
gradient will become established and intensify
during the afternoon.
This can cause distant objects
to appear elongated vertically. During the
transition between the two conditions, optical
effects will be particularly complex. In general,
therefore, gradients are likely to be less
pronounced than those described by Professor
Lehn for frozen lakes, but more complex, giving
rise to opposite conditions for distortions
in the course of a single day.
Mackal was clearly already
dissatisfied with sightings of very long necks,
and his analysis refers to the "tail,
head‑neck". It would seem, however,
that the majority of observers have not been
influenced to overstate height. The median
height for the 258 observations considered
was 0.6m (Mackal, 1976: 344)
Submerged Logs
Craig
(1982) has proposed that pine logs lying on
the loch bed may generate sufficient gas not
only to bring them to the surface, but also
to propel them before they sink again. Similarly
there have been suggestions of mats of decaying
vegetation (Burton, 1961).
It can be stated immediately that hours of television
observation of the loch floor in deepwater
have revealed no more than occasional twigs
projecting from the fine silt. If logs are
present here, they are a rarity. Intact leaves
find their way into the sediment, but at a
temperature of 5‑6 oC
decomposition is slow. No sign of gas bubbles
can be provoked by probing the sediment in
front of the camera and no gas has been observed
in cores or other mud samples brought rapidly
to the surface. Loch Ness should not be visualized
as a stagnant pond.
Two locations, however, have been noted where logs
are to be observed in quantity: off Dores
(Figure
24a,
photos 7K), where they are presumably driven
by the prevailing wind, and in Urquhart Bay
(Figure
24b, photos 12K) where in addition
they have been brought down by rivers (Figure
24c, map 7K). These logs are to be
found particularly in the shallower water
to 10m, and therefore above the summer thermocline,
but no sign of gas within them has been detected
by diving. By contrast, the leaf litter off
the River Coiltie and to a lesser extent in
other parts of the bay, does emit gas in considerable
quantity during the summer (Figure 24d,
9K). Off Fort Augustus, gas has been detected
rising over a wide area. One localised source
lay as deep as 100m. We have yet to observe
vegetation mats on the surface. Submerged
logs, however, even without gas propulsion,
may have something to answer for during underwater
photography in the seventies.
Gothic Revision
During Operation Deepscan,
Osprey low light level underwater television
(UW/TV) cameras were mounted aboard one of the cruisers to examine objects of interest
on the loch bed. Nine objects were located
by the sweeps and fixed by Decca. Anchoring
the UW/TV cruiser above loch bed contacts
200m deep, did not prove possible in the time
available and these await inspection by remote
operated submersible.
The vessel therefore
operated in shallow water to examine objects
known to be present, including the wreck of
a late‑nineteenth century sailing vessel
in Urquhart Bay. This was the area in which
controversial underwater photographs were
obtained by the Academy of Applied Science
in 1975 (Rines et al., 1976). Six photographs
were produced, interspersed with pictures
of the underside of the boat from which the
1.2 minute time lapse camera was suspended.
The camera was believed to be 40ft (12.19m)
beneath the boat, moored in 80ft (24.38m)
of water.
Initial reaction to these photographs was that
they showed the loch bed and inanimate objects
(Meredith, 1977: 6-11). More sophisticated
interpretations followed on the basis that
the camera was too far from the loch bed to
have photographed objects lying there. Furthermore,
two calibration shots of fine sediment differed
from the objects photographed. Upward pictures
of the boat, were interpreted as evidence
of agitation of the camera by possible contact
with the subject on six occasions, spread
over a period of 19 hours five minutes.
Subsequently, observations made by expedition participants
(Dinsdale, pers. comm.; and R. Raynor, pers.
comm.) have suggested that the mooring was
such as to allow the boat to swing inshore
sufficiently for the camera to touch the loch
bed.
Figure 25a (33K) constructs the implications
of the above observations and shows the probability
of the camera grounding inshore. Echo charts
(Figure
25b 10K) show the degree of variation
in depths recorded from our vessel moored
in this position and indicate objects on the
bottom. Diving inspection by Mr. R. Raynor
using an Osprey T.V.P. (underwater television
camera capable of taking still photographs
through the lens) was to reveal a number of
logs in the area, some of which are shown
(Figure
25c, 8K). One in particular, allowing
for twelve years of decay, bears considerable
resemblance to the 'gargoyle head' photograph
of 1975. Silt is also present in coarser particles
than deeper down (where the original calibration
pictures appear to have been taken) and there
is evidence of silt particles in the 1975
pictures.
Finally, the upward pictures of the boat can be
explained by the camera frame rolling or resting
on the loch bed. After the 'gargoyle head' picture, at least three consecutive
frames showed the surface; a total of 3.6
minutes, far better accounted for by the camera
lying on the bottom than by physical assault:
a pendulum motion is precluded in water.
Peaty water, combined with narrow camera angle
and small (16mm) format, were bound to render
interpretations difficult. Recognizing these
ambiguities, the naming of the Loch Ness Monster
as Nessiteras rhombopteryx (Scott and
Rines, 1975) was based upon wider evidence
and prompted by concern for conservation.
Floating Logs
Holiday (1968: 39) described
a sighting (in August 1963) which might have
come into this category had the object not
proceeded against the wind. It is recorded
that on the evening preceding the sighting,
a strong S.W. wind was blowing. Next morning
the loch was calm, after which the wind reverted
to S.W. Such conditions could have generated
a seiche. Figure
6b, showing currents and in particular
the record of physical events following a
surge (Figure
11d), makes it clear that water currents
may sometimes be in opposition to the wind
during the summer stratification. This will
be most pronounced in late summer as gradients
reach a maximum and autumn gales generate
the most powerful seiches. It is also the
time when logs and branches are washed down
by rivers. After storms, lines of logs may
be observed, drawn into the confluences of
Langmuir circulations (linear streaks of foam).
Indeed in Figure
8, showing tilting of the isotherms
by Coriolis forces, with currents in opposite
motion, it can be seen that colder water can
break surface to the left of the advancing
surface layer and move in an opposite direction.
Most students of the subject will be aware
of Adomnan's account of St. Columba's encounter
with a water beast in the River Ness. Thorpe
(1988) has pointed to another story of the
saint sailing against the wind and suggests
the above effects as an explanation.
No particular correlation with sighting reports
will be attempted here, but it would be as
well to place on record that in Loch Ness,
where physical effects are particularly significant,
logs are washed in at a time when they may
well be seen to move upwind. In strong winds
they will be disposed in straight lines associated
with foam. They may even be capable of changing
direction.
In considering alternative explanations for the
sightings record, it is noticeable that they
actually point to accuracy of observation
and vindication of quite bizarre experiences.
It would seem difficult on this basis, to
dismiss close‑range observations by
local people, who simply describe something
powerful in the loch.
2. THE SONAR CONTRIBUTION
Previous Records
Eye‑witnesses may see things they do not
understand. However, many unfamiliar observations
can be made at Loch Ness, and here of all
places it is likely that controversial interpretations
will be placed upon them. The same is true
of other forms of observation, including sonar.
The difference is that some experimental analysis
can be applied to sonar evidence, since it
seems repeatable (Mackal, 1976: 123). Furthermore,
as in surface sighting reports, it is now
possible to recognize different categories
of sonar contact. It is important to recognize
however, that a connection has yet to be demonstrated
between underwater data and what may be seen
on the surface.
Figure
26 (30K) summarizes sonar contacts
of interest gained in the sixties and seventies.
It will be seen that all results come from
sonars capable of being directed horizontally,
thus greatly increasing searching power in
mobile modes and range of coverage when used
on fixed stations.
With regard to contacts of apparently very great
dimensional extent, attention is drawn to
some difficulties arising in the interpretation
of results from fixed station work, particularly
in Urquhart Bay. The Birmingham team, after
conducting further work, were to emphasize
the refractive effects (exactly similar to
mirages) of changing thermal gradients upon
ray paths at ranges beyond 200m (Tucker and
Creasey, 1970). In this connection, Mortimer
(1973) proposed an examination of the reflective
properties of internal waves. Our observations
have shown the extent of these reflections
(Figure
11a). Whether or not the actual wave
fronts caused reflections in this case, thermal
effects in Urquhart Bay will be particularly
complex owing to seiche fronts progressively
bending into the bay as they pass. This could
be sufficient to cause horizontal refractions
in addition to vertical ones, resulting in
portions of the loch bed giving apparently
mobile returns from midwater. One such echo,
which remains stationary however, is thought
to originate from a side lobe.
Thermal and turbulent effects will be further complicated
by river outflow. In summer the river water
can be warmer than the loch, but by autumn
the opposite is the case (Figure
27a, graph 9K and
27b, echo chart 12K). The situation
is worsened by irregular loch bed contours,
existing and partially sunken moorings, considerable
boat traffic and to some extent by rising
gas. It cannot be overemphasized, however,
that Urquhart Bay is not typical of Loch Ness
as a whole.
A particular echo characteristic (again in Urquhart
Bay) reported by the Academy of Applied Science
in 1972 and 1976 consisted of sinuous multitraces
(Klein and Finkelstein, 1976). On the later
occasion, it was noted that contacts with
this signature resulted from rowing boat turbulence
(Meredith, 1977: 128‑130). During the
latter part of the 1972 trace (accompanying
the well known 'flipper' pictures) a rowing
boat was present ferrying between the vessels
engaged.
Examination of the trace shows the close‑in
returns (probably side lobe echoes) fluctuating
in range and strength, implying that the transducer
was swinging in midwater rather than resting
on the loch bed as illustrations suggest.
Some of the echoes could therefore result
from elements of topography. Lastly, the sonar
was theoretically aimed at another moored
vessel, beneath which the camera was sited.
This vessel is known to have been moored by
chain and should this have entered the beam,
it would have registered extremely strongly.
Perhaps therefore, parts of the trace consist
of this chain. The Academy recorded other
traces however, without 'turbulent' signatures,
both on fixed stations and during mobile tows.
Figure
31 (16K) shows some effects of boat
wakes on a side scan sonar record.
This leads to consideration of discrete sonar contacts
reported deep in the main water column, as
recorded by Love (1970). The biological questions
raised by contacts in this area are recognized
since, if animate, they would lie deeper than
expected for predation on scattering layer
fish and shallower than might be expected
for excursions by benthic creatures. Perplexing
observations establish however, that some
of the loch's profundal insect larvae, Sergentia
sp., occasionally migrate into
the water column in a pre‑pupal stage
and are caught within the top 30m of water.
The Project has addressed
this question since 1981, initially using
the methods of R. Love and the 'criteria for
anomaly' in terms of strength, depth and movement,
proposed by Baker and Westwood (1960).
Deep Water Contacts
In 1982, from the beginning of May to the end of
August, two scanning sonars were operated
for over 1,500 hours in day and night patrols,
mainly in the deep northern basin. The working
method is shown in Figure
28a (10K). Twelve contacts of interest
were secured by the Furuno 106A (Figure
28b, traces 33K and Figure
28c, table 12K) and a further twenty‑eight
by the Simrad SY (Figure
28d charts 64K and Figure
28e, table 22K). They appeared to
be discrete single target echoes and no other
type of contact was detected which could not
be dismissed as side echoes. Accurate navigational positioning was not possible but the
contacts appeared widely spread. On some days
none would be detected and then two or three
would result from the next patrol. Although
more contacts were gained at night with the
Simrad SY, this could be biased in that surface
conditions were then more favourable. Most
contacts occurred below the scattering layer
but sometimes strong ones appeared within
it. The search depth was limited to 160m by
side lobe echoes from the loch bed.
Strength
Clearly, contacts were
being sought with a strength greater than
that of the known fish, although perhaps in
the expectations of the 1980s, not very much
stronger. The sonars are designed to detect
large fish shoals but performed quite well
on single targets, although only through a
small proportion of the stated beam angle
to the half power point. The sensitivity of
the Furuno 106A was reduced to eliminate practically
all fish above the thermocline, which as we
have shown, are present in some abundance
over the whole loch surface. A vertical calibration
using a 20.36 cm diameter air-filled spherical
float was conducted and recorded for comparisons.
The target strength of a solid sphere is given
by:
T.S. = 10 log D2/16
(where
D is the diameter of the sphere in metres).
The calibration sphere would therefore have a strength
of -6dB. However, since the sphere was not
solid, resonance could produce considerable
inaccuracies, in general leading to a greater
strength. The actual target strength of the
sphere, when subsequently (1988) measured
in situ, with a freshwater calibrated Simrad
ES400 (38kHz) split-beam system, was -23dB.
A fairly conservative comparison can therefore
be made between the target strength of the
sphere and that of fish, using the formula
derived by Love (1971):
T.S. = 19.1
log L -0.9 log F-62
(where L is fish length in cm and F is the frequency).
For the 150kHz sonar a -26dB target would be roughly
equivalent to a fish one metre in length.
This is towards the upper limit for salmon
expected in Loch Ness.
The largest salmon reported to have been caught
in Scotland (by netting), and for which records
are available, was taken in 1891 (Mills, 1980:
4). The length was 1.35m and the weight 31.75
kg. The Loch Ness record is believed to be
23.58 kg (Witchell, 1974: 10). Figure
28c (12K) shows several contacts to
be 3dB-9dB in excess of the -26dB calibration.
Much depends upon whether a fish possesses
a swim bladder, since this accounts for over 50% of the echo strength and
also upon aspect to the beam.
With the Simrad SY, the paper recorder could not
record strength and at acceptable sensitivities,
the calibration sphere showed little reduction
in strength. Our policy was not to reduce
sensitivity beyond the point where side echoes
were readily identifiable, in case a proportion
of one should 'break through' the threshold
and give a misleading impression. We also
wished to examine weaker contacts, which could
still be of interest. Accordingly the 'calibration'
was carried out with a 30cm trout, which registered
weakly to 50m. The traces (Figure
28d, 64K), therefore include a weaker
range of strengths, some of which could be
from salmon or ferox trout. We would emphasize
that salmon would not normally be expected
to be present in midwater and are usually
fished for in the littoral zone. During the
1988 acoustic stock assessment, the maximum
fish target strengths obtained were -36dB
at 49kHz, suggesting that fish of over 30cm
are uncommon.
Some of the Simrad targets do appear strong and
obviously all represent infrequent occurrences,
but they also serve to demonstrate a 'grey
area' between the larger known fish and anything
larger still.
A last reservation concerns the nature of time-varied
gain function and scanning sonar signal processing,
which may tend to emphasize targets at the
expense of background noise. Further work
must include more precise methods of recording
target strength. The current proposal is to
use the Simrad ES400, which uses a split‑beam
principle and accurate T.V.G. to determine
target strength.
Depth
Echo‑sounding establishes
that the fish of the open water are generally
confined to the first 30m of water. Contacts
beneath this depth (about 50m range allowing
for standard tilt) were tracked for as long
as possible. Figure
29a (27K) shows the majority of strong
contacts in excess of 50m.
We consider this to be the most interesting feature
of the results. Ecologically, there seems
to be no reason why creatures in the open
water of Loch Ness should not be large, but
if that is what the contacts represent, the
depth is sometimes surprising.
Bearing in mind the difficulties associated with
horizontal beams, it was hoped that the echo‑sounder
profiles of 1983 (see map in Figure
1a,
8K), despite their low search potential, might
record the characteristic deepwater contacts.
At least the observation of fish might establish
a context. Although strength determination
is difficult, it would appear that
no contacts of the greater strengths were
acquired in 252 standard profiles, totalling
403.2 km. The rarity of moderate strength
contacts deeper than 40m in summer, is shown
in Figure
29b (16K). Situations when fish may
be observed deeper than usual are shown in
Figure 29c (21K). For example,
in May 1987 the loch showed a temperature
variation of only 6.3 C
-6.8C to a depth of 60m, and fish were detected
to at least 50m. In October 1986 contacts
were made between 80m and 120m at the southern
end of the loch. Although no temperature profile
is available, previous records suggest that
this could be in response to a lowering of
the thermocline by seiche movements. There
is some evidence of a thermocline at 60‑70m.
Similar observations have been made in previous
years. It seems difficult to shed light upon
the nature of deepwater contacts during the
intervening period, when our observations
suggest more predictable and shallower distributions.
An exceptional sequence of deepwater contacts
is shown in Figure
29d (33K). Occasional fish were noted
at depth during the fixed station work of
1984, but the range was limited to 60 metres.
It has been suggested that kelts swim deep
(Baker and Westwood, 1960), and this may be
relevant to spring observations.
Another possibility occurring to us, is that autumn
contacts could be caused by logs and branches
brought in by the rivers, although the sunken
concentrations at the leeward end of the loch
suggest that they generally float long enough
to reach there. We have seen very little evidence
of branches on the deep sediments. However,
if sinking slowly, logs could be carried considerable
distances by seiche currents.
Movement
It is known that fish do
not in general make rapid changes of depth
and this has been considered significant in
assessments of previous results (Braithwaite,
1968). We were not equipped in 1982 to record
boat movements accurately and any assessments
made in Figures
28c (12K) and 28e (22K) are restricted
to vertical estimates of target depth when
gained and lost. Margins of error are impossible
to quantify, since the effective beam width
will vary with the strength of the target
and the short tracking times achieved render
estimates tentative.
Although echo‑sounding shows very ordered
vertical movements of the fish population,
it must be understood that the trout, charr
and eels all possess 'open swimbladders' and
that rapid movements are not impossible. Occasional
records from the fixed station suggest quite
considerable vertical movements from time
to time (Figure
20d, chart 42K).
In recognition of the difficulties of judging speed
from a moving platform, a raft was four‑point
moored over 190m of water towards the
southern end of the loch in 1984 (see map
in Figure
1a, 8K). From 1st July to 24th August
the Furuno 106A was operated continuously
or every 20mins, to search the midwater around
the raft to a range of 240m and a depth of
160m. The fixed position permitted accurate
tracking and the elimination of possible side
echoes. Many moderate strength contacts (106)
were recorded to depths of approximately 40m
but not of the strengths noted in 1982. It
must be said however, that considerable equipment
problems were experienced, particularly with
the recording unit and some interesting reports
have had to be disregarded. Anchor warps obscured
and confused parts of the sweeps. Moderate
strength contacts, could now be considered
with confidence to be real single objects
and indeed to be moving. In future work it
would be important to judge movement in relation
to water currents but it should be noted that
just as sinking inanimate objects, such as
logs, may be transported by seiche currents,
so may animate ones such as fish (Figures
11a and
20c, 27K). Having considered the
contacts in relation to strength, depth and
movement, some specific alternative possibilities
should be explored.
Fish Shoals
In shallow water, Trout
have been observed to shoal on the approach
of a diver or television camera. Fish concentrate
inshore, within the scattering layer and in
autumn loose shoals are to be found at the
near surface (Figure
30, 22K). None has been observed in
deep water. Shoals often exhibit 'tails' on
echo‑sounder records, due to inter‑reflections
between the fish returning over an extended
period. Only one of our contacts showed any
vertical extent on the record.
Boat Wakes
One of our traces, gained
on shallow tilt, exhibited disproportionate
strength and had multitrace characteristics.
Inspection of the log book showed that the
contact lay in the direction of a boat passing
close by. The trace was therefore dismissed
(Figure
31, 16K).
Thermal Effects
The discrete nature of
the contacts would suggest that they are not
direct reflections from shear instabilities.
Refractions however, are quite likely to occur
as the summer progresses, although minimized
by the relatively short ranges used. Downward
refractions would cause contacts to appear
progressively shallower with increasing range.
Measurements of temperature, show the loch
to be almost isothermal from winter until
May. Thereafter, variations in depth and degree
of stratification with time and place make
predictions of long ray paths impossible.
Side Echoes
These are the most difficult
class of echoes to assess, because of the
constant proximity of the steep rocky walls.
Echoes acquired by echo‑sounder side
lobesare shown in Figure
32a (18K). With scanning sonar it
is inevitable that the horizontal beams and
side lobes must sooner or later be reflected,
to return as 'wrap around' echoes. These are
a constant and generally recognizable feature
of the traces and appear as long vertical
stripes. The short 'single target' echoes
appear distinct from these. It is conceivable
however, that localized highly reflective
rock faces at particular angles could give
rise to this form of echo.
It will be noticed
that some of the trace paths, parallel the
line of obvious side echoes as the vessel
moves towards the shoreline. Other examples
however, show range changes opposite to the
vessel's motion relative to the side walls
(Figure
32b, 13K). An experiment was mounted
in 1983 whereby the range was doubled during
a contact, thus halving the pulse repetition
frequency. This, in the case of a wrap around
echo, should result in an apparent range change.
No such changes occurred. The scanning sonar
beams depressed beyond 60 degrees recorded a 'normal' second time around echo
from the loch bed but this should be easily
recognized.
Tethered Debris
A possibility which could
not be dismissed, was that some echoes resulted
from debris in some way tethered to the loch
bed. Fishing gear jettisoned from trawlers
on passage through the Caledonian canal was
a possibility, or even lost equipment from
previous expeditions. This possibility was
explored during the 'Operation Deepscan' experiments,
described in the following supplement.
OPERATION DEEPSCAN SUPPLEMENT
Events turned full circle with the Operation Deepscan
series of 1986 and 1987 (Figure
33a, photo 10K), which were larger
scale repetitions of Dr. Baker's (1962) Cambridge
Expedition. They consisted of 'sonar curtain'
sweeps, conducted over the deep basins by
formations of echo‑sounder equipped
vessels. The objectives of Operation Deepscan
were:
1) To search the deepwater basins for contacts
of strength; to fix their positions and to
revisit the sites in order to establish whether
such contacts were tethered debris.
2) To chart objects of more general interest lying
on the loch bed. If feasible, these would
be investigated by underwater television or
subsequently examined by remote-operated submersible.
3) To continue the general scientific programme
with work in the profundal zone and observation
of the autumn fish distribution with regard
to thermal structure.
In the interests of continuity, results from 2
and 3 above have already been incorporated
into the relevant sections of this paper.
Operation Deepscan set out to overcome an impasse
which had arisen, in that contacts of strength
could be obtained by using the search and
tracking power of scanning sonars during mobile
patrols but the results were subject to the
problems of horizontal sound propagation.
Fixed station work lacked searching power,
as to a great extent did repeated echo‑sounding
profiles, since beam coverage is limited to
the area beneath the vessel.
Echo‑sounders however, yield the least ambiguous
results, since beams can be kept reasonably
clear of the side walls and penetrate thermal
gradients perpendicularly, thus minimizing
refractions. Echo-sounders also fix positions
simply and any lack of individual searching
power could be compensated by the use of so
many.
With the support of Lowrance Electronics Inc.,
vessels of the Caley Cruiser hire fleet were
equipped with X-16 sounders working on the
50kHz frequency option with 30 degree transducers
(Figure
33b, 8K). Tests carried out in October
1986 were designed to suppress mutual interference
generated by the sounders. The X-16 includes
a discrimination feature, which awaits verification
from a succession of pulses before printing
a given echo. In order to aid this facility,
the ranges around which pulse repetition frequency
changes, were staggered in sequence along
the line of vessels. Considerable success
was achieved, although at the expense of sensitivity.
Transmission interference can either be suppressed
or is easily recognizable. What is interpreted
as 'returning echo' interference (returns
from pulses initiated by other sounders),
cannot be so readily suppressed, because of
the longer length.
Figure 33c (17K), shows the effects
of operating sounders in proximity. No contacts
of interest were obtained during the 1986
tests and operations were limited by high
winds.
A fully developed experiment took place between
4th and 14th October 1987 (Figure
33d, photo 7K). From 4th to 8th October,
sweeps were conducted in the northern basin
for training purposes and to optimise equipment
settings. No contacts of interest resulted
from these.
Figure
33e (3K) shows the deployment of a
fleet used in two full sweeps of the deep
basins on 9th and 10th October. Nineteen vessels,
operating X-16 sounders as described above,
formed a line abreast at approximately 45m
spacings. Contacts gained below 30m or on
the loch bed were notified by radio to a flagship and a surface marker dropped. Follow‑up
elements, with sonar engineers aboard, then
moved in to attempt to hold contact pending
the arrival of New Atlantis. This vessel
was equipped with a Simrad EQ100 to determine
strength, a Simrad SY scanning sonar for possible
tracking, and a MK53 Decca Navigator and Racal
Decca C.V.P. 3500 plotting system to record
positions.
Figure
33f (5K) shows the coverage of the
sweeps. Sensitivity reduction resulted in
some known moderate‑strength targets
not being recorded. On the north to south
sweep of 9th October, three contacts of interest
were reported (Figure
33g, 17K). On no occasion was the
follow‑up vessel able to regain contact
in order to make strength assessments. Contact
1 (78m) may be considered of moderate strength.
Contact 2 (171m) is strong, but returning
echo interference cannot be ruled out. Contact
3 is more interesting, in that it is obviously
strong and its depth of approximately 174m
is exceptional.
Contact 3 was gained by one of the follow‑up
vessels just behind the line (the line had
failed to record the contact) to the east
of the northern basin opposite Urquhart Bay.
As the vessel stopped and turned, the contact
would appear to have crossed near the centre
of the beam. Contact was not regained by New
Atlantis but a Decca fix was taken. The
following day's sweep gained no contacts and
an extra line of five boats deployed onto
the above Decca fix also failed to regain
contact.
This is of particular interest, since at least
two fixed targets were located in the course
of the operation (see Figure
33h, 15K). Attention is drawn to the
contact of 12th October, which was gained
by a small flotilla over the western side
wall, in an area not covered by the main sweeps.
Although the follow-up was not possible until
the evening, the contact was relocated without
difficulty and so established to be fixed.
Subsequently, in July 1988, fixed contacts
recorded in the course of Operation Deepscan
were relocated in collaboration with Simrad
and investigated by the Sutec R.O.V. Sea
Owl. Both jettisoned rope and a piece
of instrumentation were in fact found. Tethered
debris has now been proved to exist in Loch
Ness but none has been found between Foyers
and Urquhart Bay where contacts were recorded
in 1982.
With regard to the interesting Contact 3, a slowly
sinking log cannot be absolutely dismissed
but failure to regain contact during the immediate
search patterns argues against this. Another
reservation must recognize the possibility
of returning echo interference. It will be
noted however, that the vessel concerned was
stationed to the flank, astern of the sweep line and transmission interference has been
eliminated from the trace. No similar trace
has been found among the other echo charts,
from both training and full sweeps, which
together represent 3,651km of search.
Conclusions
In searching the water column for previously reported
sonar contacts, scanning sonars have operated
for 1,500 hours in mobile patrols and for
over 1,000 hours on fixed stations. Echo‑sounder
records have been obtained over a distance
of 400km of standard profiles and over 4,000km
(including 1986) in massed sweeps. Contacts
of interest have been noted and a process
of elimination conducted, similar to that
applied to 'unpeeling' the scattering layers.
We suggest forms of turbulence as an explanation
for previous contacts of apparently great
dimensional extent. It is interesting that
some recognized causes of eyewitnesses' misidentification
are common to sonar, which is sensitive to
boat wakes and refractions equivalent to mirages,
while even logs are still under consideration.
Shortcomings in the work result from operational
difficulties. Mobile patrols with scanning
sonar had good searching power, but results
could be subject to ambiguities and movement
could not be convincingly demonstrated. Mooring
the station, in the interests of tracking
precision, sacrificed search potential and
the warps reduced effectiveness. Vertical
echo soundings, so desirable to reduce ambiguities,
also lacked coverage. Massed sounders bring
their own problems of interference and associated
reduction of sensitivity. Clearly, experiments
such as Operation Deepscan are limited to
a short duration; for example, only one limited
night sweep was conducted.
Work has been largely confined to the deep basins
and there remain areas where sonar methods
are more difficult. It must also be said that
if an unusual population contained a small
number of particularly large individuals,
and further, if they were not normally present
in midwater, then we might not detect them.
In the case of a single occasional migrant,
detection would be virtually impossible.
Nevertheless, contacts of interest, in terms of
strength (sometimes considerable), depth and
possible movement, do occur. By establishing
a background against which anomalies may be
judged, it is recognized that overlaps sometimes
exist in all three criteria, with the presence
and behaviour of the known fish population.
On the other hand, superficially pedestrian explanations, such as a record‑class
salmon in the main water column, deep swimming
fish shoals and midwater logs, can all be
seen to represent anomalies in themselves.
Through sonar
and underwater television the controversy
has been approached indirectly, by examination
of the environment. Even the apparently obscure
consideration of temperature is seen to dominate
not only the distribution and behaviour of
the biomass but is also a recurring element
in discussion of the controversy. Thermal
factors can have extreme effects on observations,
both above the waterline, such as mirages
and the possibilities of objects moving upwind
and also underwater, on sonar records, through
turbulence and refraction.
The habitats offered by the loch have been defined
and a framework established, bounded by the
pelagic, littoral, side wall and profundal
zones. The centre of the frame, consisting
of the bulk of the water column, contains
enigmas, which whether or not they may vindicate
sighting reports, are worthy of resolution.
In a wider sense, we are moving to fill the
data vacuum, so long the battleground of speculation
lying between protagonists in the Loch Ness
controversy.
Part C: EQUIPMENT
| Underwater Photography |
|
Osprey S.I.T. Underwater Television
Camera (O.E. 1323) |
| Resolution |
7600 T.V. lines |
| Sensitivity |
5 x 10-4 Lux |
| Lens |
5.5mm f:1.5 (corrected lens
port) |
| Focus |
Fixed 150mm to infinity |
| Angle of View |
110 degrees diagonal (in
water). |
Osprey T.V.P. (O.E. 2300/36) Capable
of taking still photographs through the
same lens. |
| Electrical Characteristics |
As above |
| Lens |
24mm f:2.5 (dome port correction) |
| Focus |
Variable 127mm to infinity |
| Angle of View |
84 degrees diagonal in water |
| Photographic Camera |
35mm format (250 shot cassette) |
| Surface Control |
Cyclops (O.E. 1210-1212) |
| Lamp |
100w variable (O.E. 1130) |
| Reproduction in this paper
by Northscene Video using Sony Videographic
Printer U.P.-811 |
Echo Sounders |
|
| Simrad EY-M |
|
| Frequency |
70kHz |
| Beam Width |
11 degrees (- 3 dB) |
| Pulse |
0.6 m/sec |
| PRF |
91 per min. at 120m range |
| TVG |
40 log R |
Simrad Skipper 603 |
Also with Kelvin-Hughes Side-scan Sonar |
| Frequency |
50kHz |
| Beam Width |
33 degrees (- 3 dB) |
| Pulse |
Variable |
| TVG |
Variable |
Lowrance Mach 1 |
192kHz 8 & 20 degree Transducers |
|
Lowrance Mach 2
|
50kHz 45degree Transducer |
Lowrance X-16 |
Dual Frequency 192kHz & 50kHz - for
Operation Deepscan : 50kHz option, 30
degree
Transducers |
Scanning Sonars |
|
Furuno F.H. 106A |
|
| Frequency |
150kHz |
| Beam Width |
6 degrees (-3dB) |
| Linked to F.C.V. colour
display for range track and tape recorder. |
| 1982 Fixed Settings |
|
| Range |
240m |
| Tilt |
Normally 30-40 degrees in
direction of travel |
| Scanning Sector |
120 degrees on automatic. Targets tracked
manually |
| Gain |
8 |
| T.V.G. Level |
3 |
| T.V.G. Time |
5 |
Simrad SY |
|
| Frequency |
80kHz |
| Beam Width |
10 degrees (-3dB) |
| Linked to AR 650 paper recorder.
Time base 54sec/cm approx. |
| 1982 Fixed Settings |
|
| Range |
250m |
| Tilt |
As above. |
| Gain |
5 (Set on AR 650 paper recorder) |
| T.V.G. |
3 + filter |
| Processing |
AGC |
Temperature Probes |
|
| Single thermistor pHOX
62T (combined oxygen temperature probe) |
| Lowrance |
Vessels |
| Motor Cruisers "New
Atlantis" available from Caley Cruisers
of Inverness. |
| Research Vessel "Simson
Echo" (Simrad demonstration vessel). |
| Motor Fishing Vessel
"Ocean Bounty". |
| Remote Operated Vehicle
Sutec Sea Owl |
Additional Equipment used in Operation
Deepscan |
| MK 53 Decca Navigator |
| Racal Decca Colour Video
Plotter CVP 3500 |
PART D: REFERENCE
Summary
Throughout
the 1980's the Loch Ness and Morar Project,
conducted volunteer expeditions to Loch Ness,
using a range of relatively simple sonar equipment.
Qualitative observations are presented through
a general description of the environment.
The elements contributing to acoustic 'scattering
layers' are analysed and separated. The role
of the temperature regime is emphasized and
turbulence due to shear instability is suggested
as a dominant cause of echoes. Initial observations
explore the relationship between physical
factors and the distribution of biomass.
1988 Loch Ness: Sonar and
Underwater Television
p195
The current neutral attitude to the Monster controversy
is made clear and discussion is mainly against
the accumulated background of physical and
biological information. A particular study
has been made of strong deepwater sonar contacts,
reported by the Loch Ness Investigation Bureau
of the 1960s. The presence of occasional contacts,
which are exceptional in terms of strength
and depth is established, while the extent
to which they represent anomalies is discussed.
Although these may not meet popular expectations
and whatever their relationship to the controversy
may be, they would seem to merit further enquiry.
Acknowledgements
The information presented in this paper resulted
from the perseverance and discipline of the
many individual volunteers who made up the
field membership of the Loch Ness and Morar
Project throughout the 1980's. The following
organizations also provided contingents: St.
Benedict's Abbey Combined Cadet Forces, the
Sea Cadet Organization, the Venture Scout
Movement, the Fort George Volunteers, the
West Yorkshire Fire Services Sailing Club,
the Royal Corps of Transport Sailing Club
(42 Sqd.), the Drake Fellowship and the Dockland
Scout Project.
The following companies were generous with the
loan of equipment: Osprey Electronics, Simrad
Albatross Ltd., Lowrance Electronics Inc.,
Tamtech Ltd., Chloride U.K. Ltd., Swiftech
Ltd., Sutec U.K. Ltd. and Instrument Rentals
U.K. Ltd.
Special thanks are due to Mr. and Mrs J. Hogan
and Caley Cruisers Ltd. for the constant support
they gave with elements of their hire fleet,
particularly the New Atlantis sonar-equipped
motor cruiser.
The scientific programme was undertaken in collaboration
with the following organizations: the Ness
District Salmon Fisheries Board and the Department
of Zoology, Royal Holloway College. The British
Ecological Society kindly provided financial
support for work on pelagic fish.
Valuable support and advice was given by the following
individuals: Dr. H.J.B. Birks (cores), Dr.
A. Duncan (zooplankton), Dr. J. Evans (phytoplankton),
Mr. R. Greer (Charr), Mr. J.D. Hamilton (sediments),
Dr. T. Lindem (acoustic fish stock survey),
Fr. Andrew McFillop (weather records), Dr.
S.A. Thorpe (physical limnology), Mr. P. Wilkinson
(fish), and Dr. B. Woodward (acoustics).
We also wish to thank the
residents of Drumnadrochit, Fort Augustus,
and Dores for all their goodwill and support,
especially Mrs M.Gore, the Hon. J. Kirkwood,
Mr. A. Menzies, Mr.G. Menzies, Mr. A. Harmsworth
and Mr. R.A. Bremner.
1988 The Scottish Naturalist
p196
General
support was received from the Loch Ness Centre
and the Highlands and Islands Development
Board. Reproduction of videotape
pictures are by North Scene Video. Photomicrographs
are by Mr. C.J. Chesney. We are extremely
grateful to Mrs Jane C. Shine for all the
other illustrations and Deepscan photographs.
References
ANON. (1930). What was it? A strange experience on Loch Ness.
Northern Chronicle, 27th August 1930.
BAILEY-WATTS, A.E. and DUNCAN, P. (1981). The phytoplankton.
In The Ecology of Scotland's Largest Lochs:
Lomond, Awe, Ness, Morar and Shiel. (Ed. P.S.
Maitland). Monographiae Biologicae, 44: 91-118.
The Hague: Junk.
BAKER, P.F. (1962). Cambridge University Loch Ness Expedition
Report. Cambridge.
BAKER, P.F. and WESTWOOD, M. (1960). Underwater detective work.
Scotsman, 14th September 1960.
BAUER, H.H. (1986). The Enigma of Loch Ness. Urbana and Chicago:
University of Illinois Press.
BINNS, R. (1983). The Loch Ness Mystery Solved. Shepton Mallet
(Somerset): open Books.
BRAITHWAITE, H. (1968). Sonar
picks up stirrings in Loch Ness. New Scientist
(19th December 1968), 40: 664-666.
BRUNDIN, L.
(1949). Chironomiden und andere Bodentiere
der Sudschweden Urgebirsseen. Report of the
Institute of Freshwater Research [Drottningholm],
30: 1-914.
BURTON,
M. (1961). The Elusive Monster. London: Hart-Davies.
CAMPBELL,
S. (1986). The Loch Ness Monster. The Evidence.
Wellingborough (Northampton): Aquarian Press.
CRAIG, R.P. (1982). Loch Ness: the Monster unveiled. New Scientist
(5th August 1982), 95: 354-357.
CRAWFORD, G.I. (1937). An amphipod Eucrangonyx gracilis s.i.
Smith, new to Britain. Nature, 139: 327.
DINSDALE,
T. (1961). Loch Ness Monster. London: Routledge
and Kegan Paul, 1961; Philadelphia: Chilton,
1962; 2nd ed., Routledge and Kegan Paul, 1972;
3rd ed., 1976; 4th ed., 1982.
EASTAUGH,
R.W. (1970). Report of freshwater trials of
the submarine Pisces. Loch Ness Investigation,
Annual Report, 1969: 7-8.
FITTER, R.S.R. (1988). The Loch Ness Monster: Saint
Columba to the Loch Ness Investigation Bureau.
Scottish Naturalist,1988: 43‑54.1
1988 Loch Ness: Sonar and Underwater
Television p197
GOULD R.T.
(1934). The Loch Ness Monster and Others.
London: Geoffrey Bles, 1934; New York: University
Books Inc., 1969; New Jersey: Citadel Press,
1976.
GREENLAW, C.F. (1979). Acoustical estimation of
zooplankton populations. Limnology and Oceanography,
24: 226‑242.
HARDEN‑JONES, F.R. (1969). Fish Migration. London: Edward,
Arnold.
HOLIDAY, F.W.
(1968). The Great Orm of Loch Ness. London:
Faber and Faber, 1968; Toronto: George J.
McLeod, 1969; New York: W.W. Norton, 1969;
New York: Avon, 1970.
HUNTER, W.D.R.
and SLACK, H.D. (1958). Pisidium conventus
Clessin in. Loch Lomond. Journal of Conchology,
24: 245-247.
KLEIN, M.
and FINKELSTEIN, C. (1976). Sonar serendipity
in Loch Ness. Technology Review, December
1976: 44-57.
LEE, G.W.
and COLLET, L.W. (1908). Notes on the deposits
of Loch Ness. Geographical Journal, 31: 58-61.
LEHN, W.H. (1979). Atmospheric refraction and lake monsters.
Science, 205: 183-185.
LOVE, R. (1970). Sonar results from Loch Ness. Loch Ness Investigation,
Annual Report, 1969: 9-12.
LOVE, R.H.
(1971). Dorsal aspect target strength of individual
fish. Journal of the Acoustical Society of
America, 49: 816-823.
MACKAL,
R.P. (1976). The Monsters of Loch Ness. Chicago:
Swallow, 1976; London: Macdonald and Janes's,
1976; London: Futura, 1976.
MACKAL, R.P.
and LOVE, R. (1970). Chemical sampling in
Loch Ness. Loch Ness Investigation, Annual
Report, 1969: Appendix iii, p.18.
MAITLAND,
P.S. (1977). Freshwater fish in Scotland in
the 18th, 19th and 20th centuries. Biological
Conservation, 12: 265-278.
MAITLAND,
P.S. (Ed.) (1981). The Ecology of Scotland's
Largest Lochs: Lomond, Awe, Ness, Morar and
Shiel. Monographiae Biologicae, Vol. 44. The
Hague: Junk.
MEREDITH, D. (1977). The Search at Loch Ness. New York: Quadrangle
(New York Times).
MILLS, D. (1980). Scotland's King of Fish. Edinburgh: Blackwood.
MORTIMER,
C.H. (1952). Water movements in lakes during
summer stratification; evidence from the distribution
of temperature in Windermere. Philosophical
Transactions of the Royal Society of London,
Series B, 236: 355-404.
MORTIMER,
C.H. (1955). Some effects of the Earth's rotation
on water movement in stratified lakes. Proceedings
of the International Association of Applied
Limnology, 12: 66-77.
1988 The Scottish Naturalist p198
MORTIMER, C.H. (1973).
The Loch Ness Monster - limnology or paralimnology?
Limnology and Oceanography, 18: 343‑345.
MURRAY, J. (1904). Biology of Loch Ness. Geographical Journal,
24: 442-443.
MURRAY, J. and PULLAR, L. (Editors). (1903-08). Bathymetrical survey
of the fresh‑water lochs of Scotland.
Geographical Journal, Vols. 22-31. A series
of papers by various contributors, preliminary
to the six volume publication of 1910.
MURRAY, J.
and PULLAR, L. (1908a). Bathymetrical survey
of the fresh‑water lochs of Scotland.
Part XIII -Lochs of the Ness basin. Geographical
Journal, 31: 42-72.
MURRAY, J. and PULLAR, L. (1908b). Mirages on Loch Ness. Geographical
Journal, 31: 61-62.
MURRAY, J.
and PULLAR, L. (Editors). (1910). Bathymetrical
Survey of the Scottish Fresh-Water Lochs.
Vols. 1-6. Edinburgh: Challenger office.
NORTHCOTE,
T.G. (1964). Use of a high frequency sounder
to record distribution and migration of Chaoborus
larvae. Limnology and Oceanography, 9: 87-91.
RAZDAN, R. and KIELAR, A. (1984).
Sonar and photographic searches for the Loch
Ness Monster: a reassessment. Skeptical Inquirer,
9(2): 147-158.
REYNOLDSON,
T.B., SMITH, B.D. and MAITLAND, P.S. (1981).
A species of North American triclad (Paludicola;
Turbellaria) new to Britain found in Loch
Ness, Scotland. Journal of Zoology, 193: 531-539.
RINES, R.H.,
WYCKOFF, C.W., EDGERTON, H.E. and KLEIN, M.
(1976). Search for the Loch Ness Monster.
Technology Review, March/April 1976: 25-40.
RYDER, R.A.
(1964). Chemical characteristics of Ontario
lakes, with reference to a method for estimating
fish production. Ontario Department of Lands
and Forests, Section Report (Fish), 48: 75pp.
RYDER, R.A.
(1965). A method for estimating the potential
fish production of north‑temperate lakes.
Transactions of the American Fisheries Society,
94: 214-218.
SCHEIDER,
W. and WALLIS, P. (1973). An alternate method
of calculating the population density of Monsters
in Loch Ness. Limnology and Oceanography,
18: 343.
SCHRODER,
R. (1962). Vertikalverteilung des Zooplanktons
und Thermokline. Archiv fur Hydrobiologie,
Supplement-Bd., 25: 401-410.
SCOTT, P. and RINES, R.H. (1975). Naming the Loch Ness Monster.
Nature, 258: 466-468.
SHELDON, R.W.
and KERR, S.R. (1972). The population density
of Monsters in Loch Ness. Limnology and Oceanography,
17: 796-798.
SHINE,
A.J. (1976). Loch Morar Expedition 76 Report.
1988 Loch Ness: Sonar and Underwater
Television p199
SIMPSON, J.H.
(1970). Temperature microstructure of Loch
Ness. Proceedings of the Challenger Society,
4: 64.
SMITH, I.R.
and LYLE, A.A. (1979). Distribution of Freshwaters
in Great Britain. Edinburgh: Institute of
Terrestrial Ecology.
THORPE, S.A.
(1974). Evidence of Kelvin‑Helmholtz
billows in Loch Ness. Limnology and Oceanography,
19: 973-976.
THORPE, S.A.
(1977). Turbulence and mixing in a Scottish
loch. Philosophical Transactions of the Royal
Society of London, Series A, 286: 125-181.
THORPE, S.A.
(1988). "...light on obscure oceanographical
problems"; an historical review of studies
of the physics of Loch Ness. Scottish Naturalist,
1988: 17-33.
THORPE, S.A., HALL, A. and CROFTS, 1. (1972). The internal
surge in Loch Ness. Nature, 237: 96~98.
TUCKER, D.G.
and CREASEY, D.J. (1970). Some sonar observations
in Loch Ness. Proceedings of the Challenger
Society, 4: 91-92.
WALKER, A.F.,
GREER, R.B. and GARDNER, A.S. (1988). Two
ecologically distinct forms of Arctic Charr,
Salvelinus alpinus (L.) in Loch Rannoch, Scotland.
Biological Conservation, 43: 43-61.
WATSON, E.R.
(1904). Movements of the waters of Loch Ness,
as indicated by temperature observations.
Geographical Journal, 24: 430-437.
WEDDERBURN, E.M. (1907). The temperature of the
fresh‑water lochs of Scotland, with
special reference to Loch Ness. With appendix
containing observations made in Loch Ness
by members of the Scottish Lake Survey. Transactions
of the Royal Society of Edinburgh, 45: 407-489.
WILLIAMSON, G.R. (1987). Seals in Loch Ness. Inverness: Privately
published report.
WITCHELL, N. (1974). The Loch Ness Story. Lavenham
(Suffolk) Terence Dalton,
1974; 2nd ed., 1976 (London Book Club Associates,
1979); rev. ed., Harmondsworth: Penguin, 1975;
rev. ed., London: Corgi, 1982.
WOODS, J.D.
(1968). Wave-induced shear instability in
the summer thermocline. Journal of Fluid Mechanics,
32: 791-800.
Mr. Adrian
J. Shine and Mr. David S. Martin,
Loch Ness and Morar Project,
Loch Ness Centre,
DRUMNADROCHIT, Inverness‑shire
IV63 6TU.
Copyright:
May be used for private research only
|

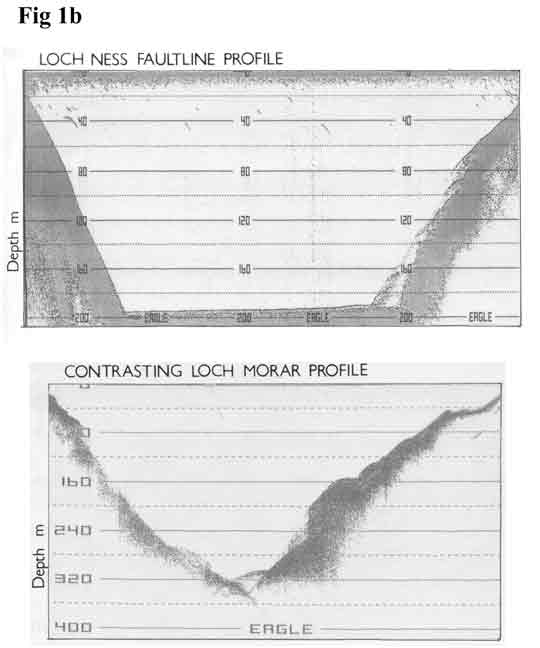{kind=link}
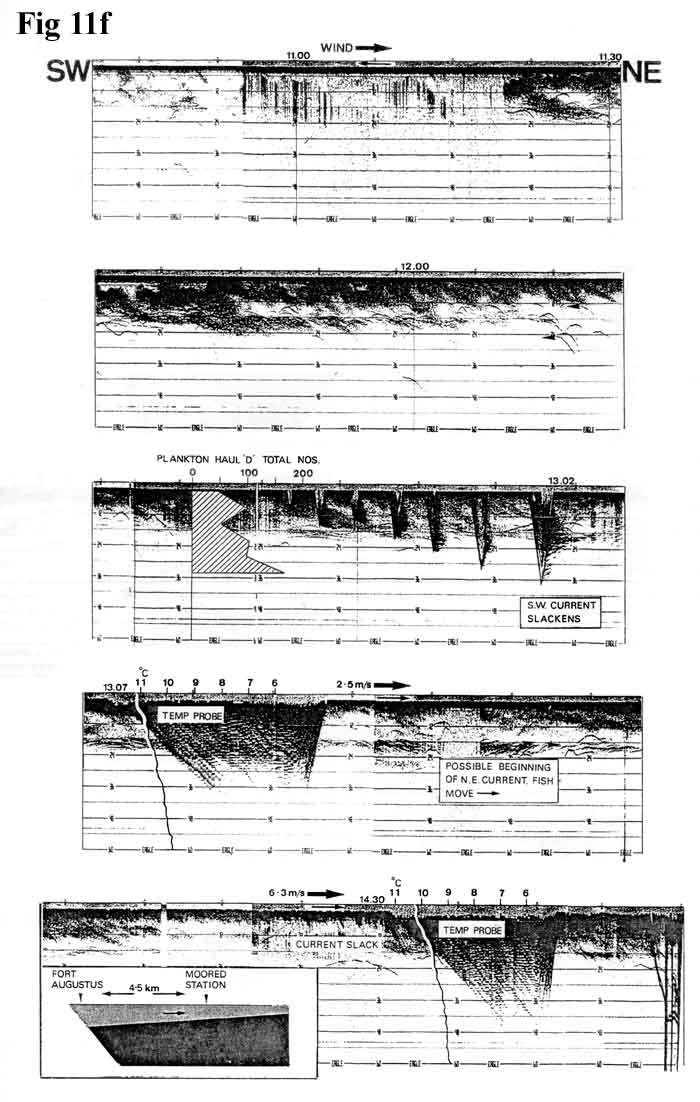{kind=link}
{kind=link}
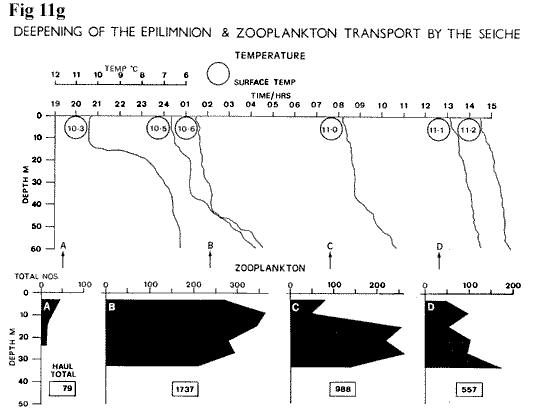{kind=link}
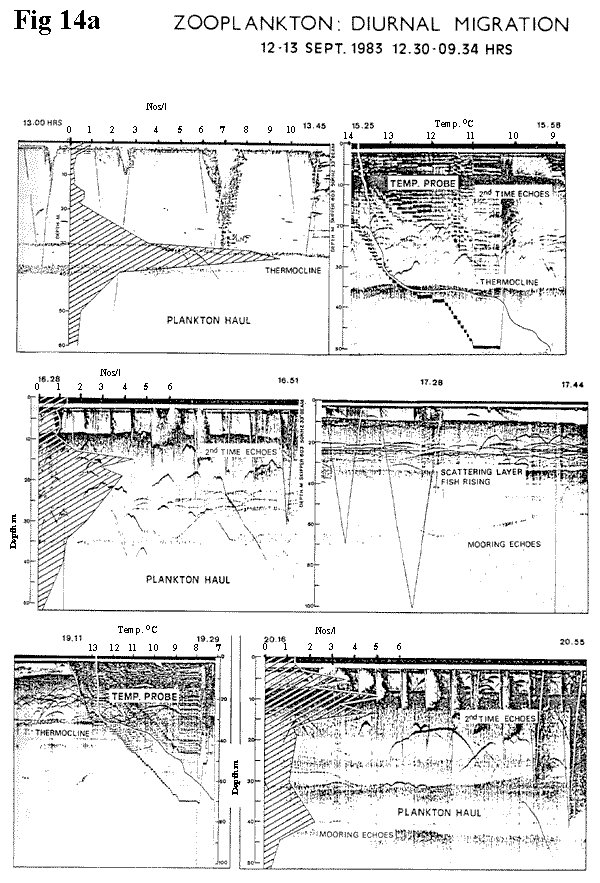{kind=link}
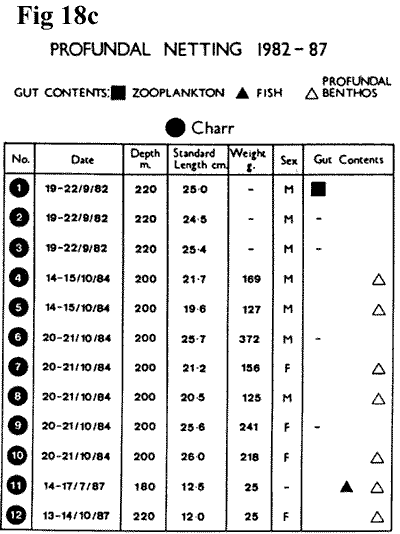{kind=link}
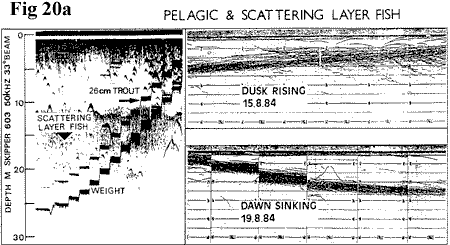{kind=link}
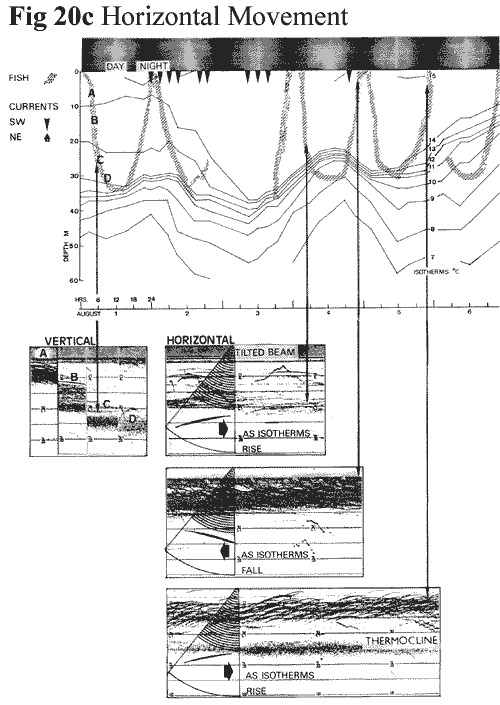{kind=link}
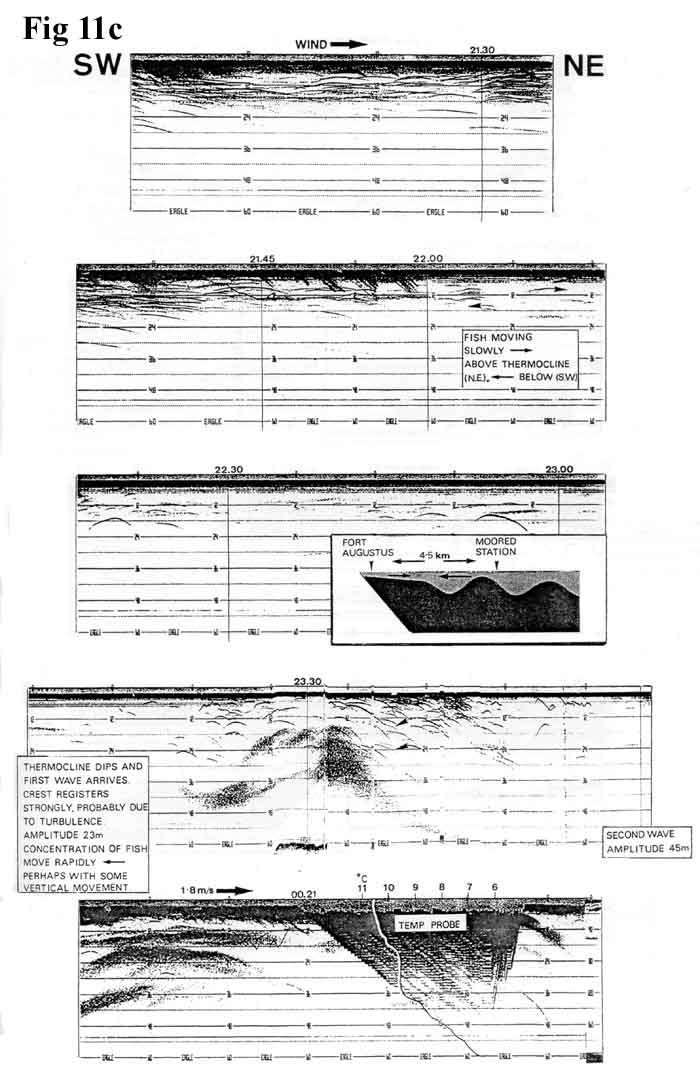{kind=link}
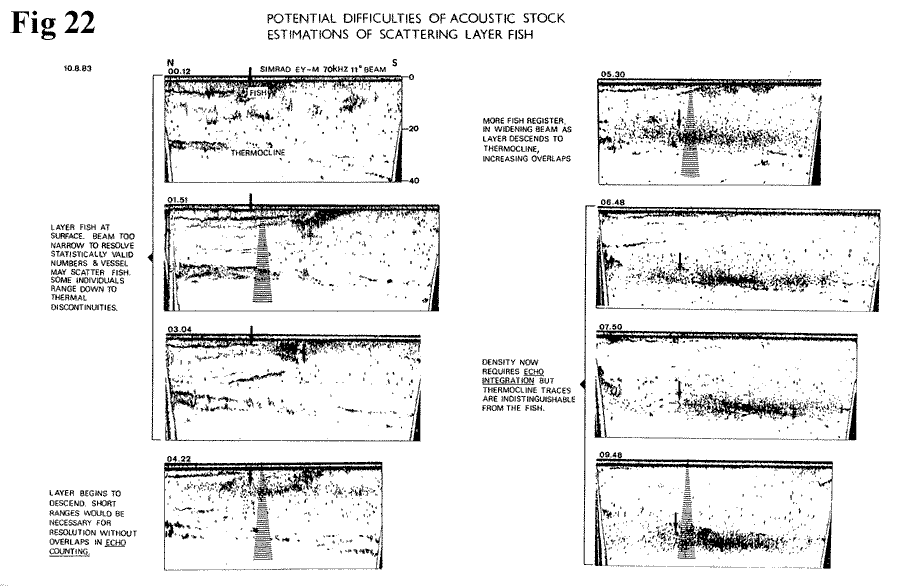{kind=link}
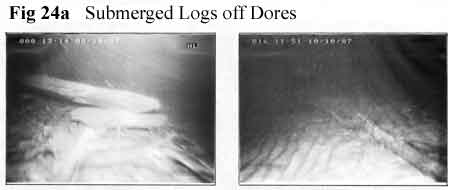{kind=link}
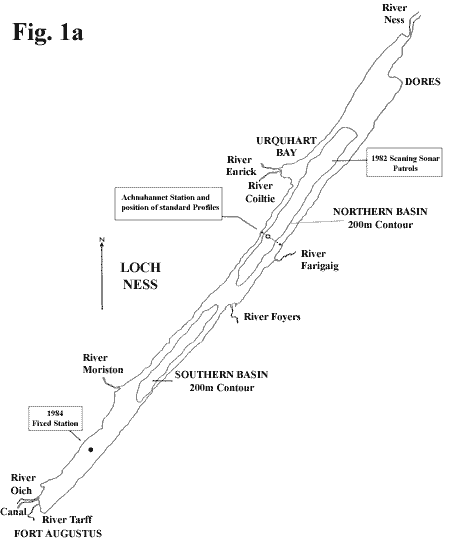{kind=link}